
 Início
InícioÚltimos assuntos
Tópicos mais visitados
Tópicos mais ativos
SEU IP


Protegendo a integridade do genoma através do reparo extraordinário do DNA
Página 1 de 1

24042011
Protegendo a integridade do genoma através do reparo extraordinário do DNA
Protegendo a integridade do genoma através do reparo extraordinário do DNA
Sábado, Abril 23, 2011
Safeguarding Genome Integrity Through Extraordinary DNA Repair
Berkeley Lab Scientists Discover Dynamic Double-Strand Break Repair in Heterochromatin
APRIL 18, 2011
Paul Preuss 510-486-6249 paul_preuss@lbl.gov
DNA is under constant attack, from internal factors like free radicals and external ones like ionizing radiation. About 10 double-strand breaks – the kind that snap both backbones of the double helix – occur every time a human cell divides. To prevent not only gene mutations but broken chromosomes and chromosomal abnormalities known to cause cancer, infertility, and other diseases in humans, prompt, precise DNA repair is essential.
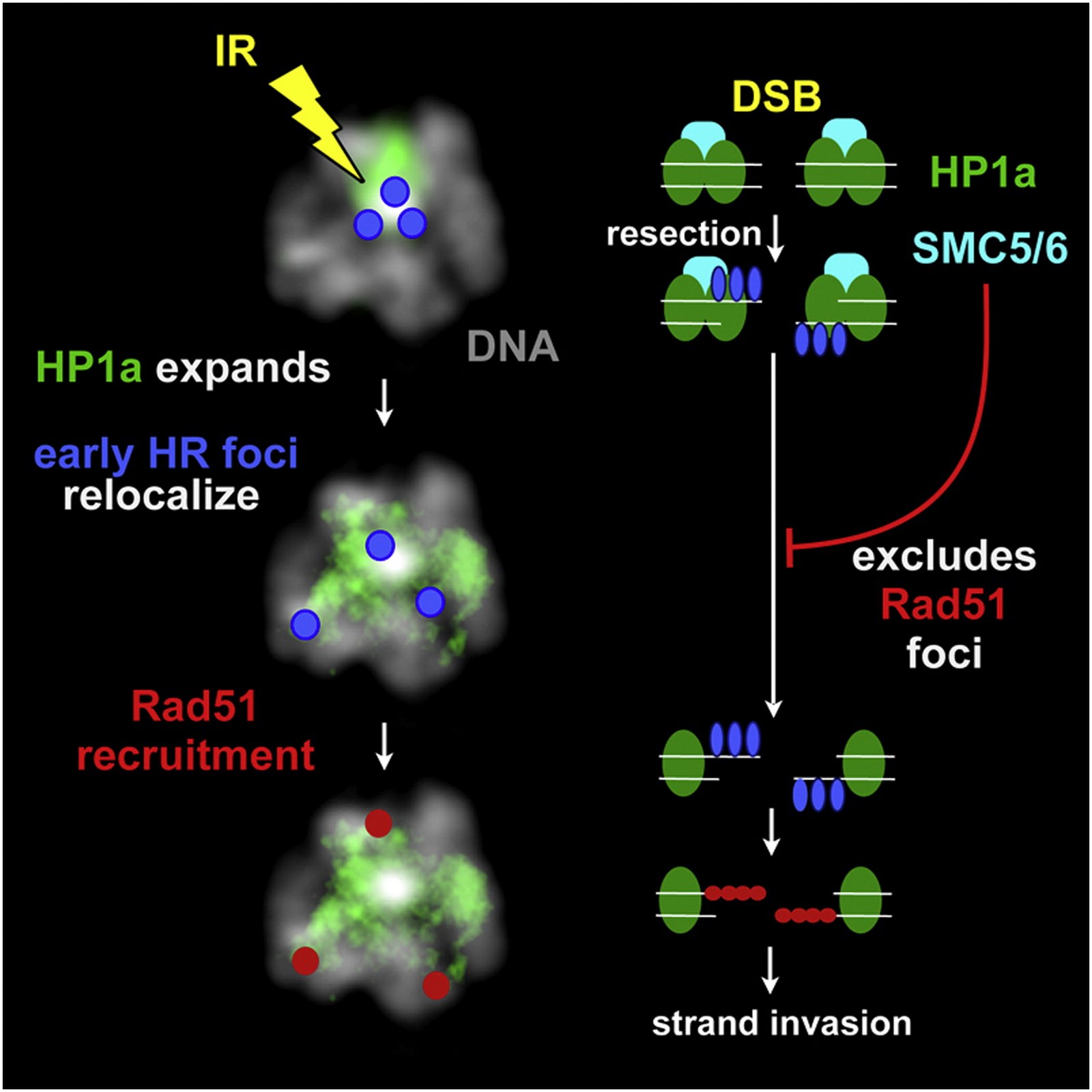
Scientists at the U.S. Department of Energy’s Lawrence Berkeley National Laboratory (Berkeley Lab), working with cell lines of the fruit fly Drosophila melanogaster, have discovered an unsuspected and dramatic process by which double-strand breaks in heterochromatin – one of the two major kinds of chromatin that make up chromosomes, which accounts for a third of the chromatin in both humans and fruit flies – are repaired in a series of steps. The repair starts where the break occurs, but stalls until the repair site physically moves away from the original heterochromatin region, before continuing to completion.
Unlike euchromatin, where most of an organism’s genes reside and where most DNA consists of long, unrepetitive sequences of base pairs, DNA in heterochromatin consists mostly of short repeated sequences that don’t code for proteins; indeed, heterochromatin was long regarded as containing mostly “junk” DNA.

Heterochromatin (purple) accounts for a third of the chromatin in both humans and fruit flies. Some heterochromatin forms the telomeres that cap the ends of the chromatids, and much is concentrated near the centromere, where sister chromatids are joined. Accurate repair of double-strand breaks in heterochromatin is challenging, because most of its DNA consists of short, repeated sequences.
Heterochromatin is now known to be anything but junk, playing a crucial role in organizing chromosomes and maintaining their integrity during cell division. It is concentrated near centromeres, where chromatids are in closest contact, which are required to transmit chromosomes from one generation to the next. Maintaining heterochromatin structure is necessary to the normal growth and functions of cells and organisms.
“Heterochromatin poses more of a problem for DNA repair than euchromatin,” says Gary Karpen, whose group in Berkeley Lab’s Life Sciences Division discovered the new repair mechanism. “It has lots of short sequences – many of them only about five base-pairs each – which are repeated millions of times.”
“Repair of simple repeated sequences is particularly challenging,” says Irene Chiolo, first author of the group’s paper reporting the results in the journal Cell. “They can promote chromosome aberrations, with severe consequences for the genome stability of dividing cells” – abnormalities that are a hallmark of cancer cells and cause birth defects.
Finding the right path
With the stakes so high, how can cells insure fast, accurate repair of double-strand breaks? Two main repair pathways are available. One method, nonhomologous end-joining, simply cleans up the ends of the broken strands and glues them back together regardless of sequence. This might seem a good choice for heterochromatin: it almost always creates small deletions or mutations, but these are in repetitive, noncoding sequences and do not affect genes.
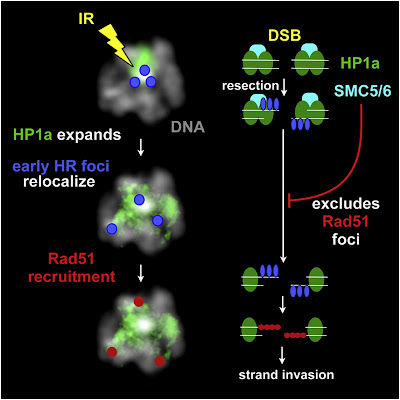
Far more accurate but more complex is homologous recombination, a mechanism involving many steps where something could go wrong. Upon detecting a double-strand break in DNA, several proteins rush to the damaged area. The protein machinery trims back the ends of the broken strands (called ‘resection’) to produce single-strand regions recognized by other proteins, including one called ATRIP.
Another protein, Rad51, is recruited to form filaments on the single-stranded DNA. Rad51 and its associated proteins search for a complementary sequence of DNA in a neighboring chromatid or homologous chromosome. They invade and open that DNA to form a “D-loop” – like untwisting a rope to open and expose its individual strands. Using the exposed complementary sequence as a template, proteins rebuild the broken DNA into a copy of the sequence that was originally damaged; in this way the broken double strand is remade with its damaged section accurately reproduced.
It’s an ideal method for repairing breaks in gene-rich euchromatin. In repetitive heterochromatin, however, danger arises because completely different chromosomes lying close to the site of the break may have great lengths of repeated short sequences that look identical to the region around the break itself. What starts as a repair process may end up splicing different chromosomes together, a common abnormality in cancer cells.
For heterochromatin to employ such a potentially risky repair process seemed counterintuitive. In earlier experiments looking for key signs of repair in mouse heterochromatin after irradiation, classic markers of double-strand break repair by either nonhomologous end-joining or homologous recombination were both absent. In fact it seemed possible that, somehow, such breaks didn’t occur in heterochromatin.
“There were no signs of repair half an hour after the cells were exposed to ionizing radiation,” says Karpen. “But our group looked at Drosophila cells just 10 minutes after radiation exposure. Now the early signs of homologous recombination were clearly evident.”
After half an hour, however, these signs too – signals from modified histones, the component proteins that form the “spools” around which the DNA “thread” is wound in chromatin, as well as signals from ATRIP recruitment – were missing from the heterochromatin domain. What had become of the repair process?
...
Read more here/Leia mais aqui: Berkeley Lab News Center
+++++
Cell
Volume 144, Issue 5, 4 March 2011, Pages 732-744
Double-Strand Breaks in Heterochromatin Move Outside of a Dynamic HP1a Domain to Complete Recombinational Repair
Irene Chiolo1, Aki Minoda1, Serafin U. Colmenares1, Aris Polyzos1, Sylvain V. Costes1 and Gary H. Karpen1, 2,
1 Department of Genome Dynamics, Lawrence Berkeley National Laboratory, Berkeley, CA 94720, USA
2 Department of Molecular and Cell Biology, University of California, Berkeley, Berkeley, CA 94720, USA
Received 3 June 2010;
revised 15 December 2010;
accepted 7 February 2011.
Published online: February 24, 2011.
Available online 25 February 2011.
Summary
Double-strand breaks (DSBs) in heterochromatic repetitive DNAs pose significant threats to genome integrity, but information about how such lesions are processed and repaired is sparse. We observe dramatic expansion and dynamic protrusions of the heterochromatin domain in response to ionizing radiation (IR) in Drosophila cells. We also find that heterochromatic DSBs are repaired by homologous recombination (HR) but with striking differences from euchromatin. Proteins involved in early HR events (resection) are rapidly recruited to DSBs within heterochromatin. In contrast, Rad51, which mediates strand invasion, only associates with DSBs that relocalize outside of the domain. Heterochromatin expansion and relocalization of foci require checkpoint and resection proteins. Finally, the Smc5/6 complex is enriched in heterochromatin and is required to exclude Rad51 from the domain and prevent abnormal recombination. We propose that the spatial and temporal control of DSB repair in heterochromatin safeguards genome stability by preventing aberrant exchanges between repeats.
Graphical Abstract
Highlights
► DSB detection and processing occur very efficiently in heterochromatin after IR ► Heterochromatic DSBs move outside the expanding HP1a domain to complete HR repair ► HP1a expansion and DSB relocalization require checkpoint and resection proteins ► Smc5/6 prevent Rad51 foci formation and aberrant recombination within the HP1a domain
+++++
Professores, pesquisadores e alunos de universidades públicas e privadas com acesso ao site CAPES/Periódicos podem ler gratuitamente este artigo da Cell e de mais 22.440 publicações científicas.
+++++
PERGUNTAS INDISCRETAS DESTE BLOGGER:
Excerto do artigo do Berkeley Lab News Center:
"To prevent not only gene mutations but broken chromosomes and chromosomal abnormalities known to cause cancer, infertility, and other diseases in humans, prompt, precise DNA repair is essential."
"Homologous recombination is a complex mechanism with multiple steps, but also with many points of regulation to insure accurate recombination at every stage. This could be why this method has been favored during evolution. The machinery that relocalizes the damaged DNA before loading Rad51 might have evolved because the consequences of not having it would be terrible."
A Nomenklatura científica uma hora dessas deve estar pedindo a cabeça do Paul Press. Razão? A definição de evolução dele contraria a definição consensual aceita pela Akademia: um processo de mero acaso, fortuita necessidade ao longo do tempo filtrado pela seleção natural ou n mecanismos evolucionários. Repare bem:
Sendo um processo ao acaso, a evolução não tem objetivo e nenhum propósito, e nem se importaria se algo surgisse ou não.
1. Então, alguém me explique aqui, como que um processo sem inteligência pode favorecer processo tão complexo como esse?
2. Como que um processo sem inteligência pode saber que as consequências de não ter este processo complexo seria terrível para as formas biológicas?
3. E esta última para pensar cum grano salis, COMO isso [um processo cego, aleatório, não guiado e que não tem em mente qualquer finbalidade nas formas biológicas] motivou uma maquinaria e mecanismos tão complexos para evitar consequências terríveis?
Gente, a linguagem é TELEOLÓGICA, e isso é defendido pelos teóricos e proponentes da teoria do Design Inteligente!!!
Fui, nem sei por que, pensando que a cada dia que passa com os avanços e descobertas da ciência, Darwin cada vez mais vai sumindo de cena ou então a Nomenklatura científica se vê forçada a elaborar verdadeiros epiciclos epistêmicos a la Ptolomeu, teorias ad hoc, para salvar a cara de Darwin, o homem que teve a maior ideia que toda a humanidade já teve!!!
A nova teoria geral da evolução - a SÍNTESE EVOLUTIVA AMPLIADA vai ter que considerar tudo isso.

Eduardo- Mensagens : 5997
Idade : 54
Inscrição : 08/05/2010
Compartilhar este artigo em:
Protegendo a integridade do genoma através do reparo extraordinário do DNA :: Comentários
Nenhum comentário
» A integridade científica em risco diante da comercialização da ciência
» RNA "lixo": de mensageiro a guardião da integridade genômica - mero acaso, fortuita necessidade ou design inteligente???
» A Semana Através dos Tempos
» A flor do genoma
» O domínio da mente no cristianismo através da música
» RNA "lixo": de mensageiro a guardião da integridade genômica - mero acaso, fortuita necessidade ou design inteligente???
» A Semana Através dos Tempos
» A flor do genoma
» O domínio da mente no cristianismo através da música
Permissões neste sub-fórum
Não podes responder a tópicos







 Seja fã Forumeiros
Seja fã Forumeiros
» Acordem adventistas...
» O que Vestir Para Ir à Igreja?
» Ir para o céu?
» Chat do Forum
» TV Novo Tempo...
» Lutas de MMA são usadas como estratégia por Igreja Evangélica para atrair mais fiéis
» Lew Wallace, autor do célebre livro «Ben-Hur», converteu-se quando o escrevia
» Ex-pastor evangélico é batizado no Pará
» Citações de Ellen White sobre a Vida em Outros Planetas Não Caídos em Pecado
» Viagem ao Sobrenatural - Roger Morneau
» As aparições de Jesus após sua morte não poderiam ter sido alucinações?