
 Início
InícioÚltimos assuntos
Tópicos mais visitados
Tópicos mais ativos
SEU IP


A Tese da Complexidade Irredutível
Página 1 de 1

01042010
A Tese da Complexidade Irredutível

Complexidade irremediável na Science?
Quinta-feira, Novembro 11, 2010
Science 12 November 2010:
Vol. 330. no. 6006, pp. 920 - 921
DOI: 10.1126/science.1198594
PERSPECTIVES
CELL BIOLOGY:
Image not related to this article/Imagem não relacionada a este artigo
Irremediable Complexity?
Michael W. Gray,1 Julius Luke,2 John M. Archibald,1 Patrick J. Keeling,3 W. Ford Doolittle1
Many of the cell's macromolecular machines appear gratuitously complex, comprising more components than their basic functions seem to demand. How can we make sense of this complexity in the light of evolution? One possibility is a neutral ratchet-like process described more than a decade ago (1), subsequently called constructive neutral evolution (2). This model provides an explanatory counterpoint to the selectionist or adaptationist views that pervade molecular biology (3).
1 Department of Biochemistry and Molecular Biology, Dalhousie University, Halifax, Nova Scotia B3H 1X5, Canada.
2 Institute of Parasitology, Czech Academy of Sciences and Faculty of Sciences, University of South Bohemia, 37005 eské Budjovice, Czech Republic.
3 Department of Botany, University of British Columbia, Vancouver, British Columbia V6T 1Z4, Canada.
E-mail: ford@dal.ca
+++++
Professores, pesquisadores e alunos de universidades públicas e privadas com acesso ao site CAPES/Periódicos podem ler gratuitamente este artigo da Science e de mais 22.440 publicações científicas.
+++++
NOTA DESTE BLOGGER:
Ué, mas complexidade irredutível de sistemas biológicos não é pseudociência? Criacionismo disfarçado? Coisa de quem não sabe o que é e nem como se faz ciência? Ora, convenhamos, complexidade é coisa dos teóricos e proponentes da teoria do Design Inteligente. Quer dizer então que há plausibilidade científica nesta nova teoria???
Fui, nem sei por que, pensando que mais dia menos dia a Nomenklatura científica vai ter que abrir espaço para artigos assim como este acima.
Complexidade Irredutível refutada?
Estudo refuta a complexidade irredutível
Pesquisa mostra como seleção natural reusa peças para criar novas máquinas moleculares Kenneth Chang escreve para o New York Times:
COMENTÁRIO SOBRE A NOTÍCIA FEITO PELO COORDENADOR DO NBDI (NÚCLEO BRASILEIRO DE DESIGN INTELIGENTE)Reconstruindo genes de animais extintos, cientistas demonstraram pela primeira vez o processo gradual pelo qual a evolução cria uma nova peça de maquinário molecular ao reusar e modificar partes existentes.Os pesquisadores dizem que sua descoberta, publicada na última sexta-feira no periódico Science, dá um contra-argumento aos céticos da evolução, os quais questionam como uma progressão de pequenas mudanças poderia produzir os intricados mecanismos encontrados nas células vivas.
A evolução da complexidade é uma velha questão da biologia evolutiva, disse Joseph Thornton, professor de biologia na Universidade do Oregon e autor principal do artigo científico. Não há controvérsia sobre se esse sistema evoluiu. A questão para os cientistas é como ele evoluiu, e foi isso o que o nosso estudo mostrou. Os experimentos de Thornton miraram em dois receptores de hormônios. Um é um componente de sistemas de resposta a estresse. O outro, apesar de ter uma forma parecida, participa de processos biológicos diferentes, incluindo função renal em animais.
Hormônios e seus receptores funcionam como pares de chave e fechadura. Seu encaixe levou à questão de como novos pares evoluem, já que um sem o outro aparentemente seria inútil. Os pesquisadores encontraram os equivalentes modernos ao receptor de hormônio de estresse na lampréia, um peixe primitivo, e também em arraias.
Depois de comparar semelhanças e diferenças nos genes, os cientistas concluíram que ele descendia de um gene ancestral comum há 450 milhões de anos - antes de os animais conquistarem terra firme.O grupo, então, recriou o receptor ancestral em laboratório, e descobriu que ele se encaixava no hormônio de estresse, o cortisol, e no de regulação renal, a aldosterona.Assim, o receptor para a aldosterona existia antes da aldosterona aparecer - o hormônio só evoluiu em animais terrestres. Ele tinha uma função diferente e foi aproveitado para participar de um novo sistema complexo quando o hormônio apareceu, disse Thornton. Ele afirma que o estudo refuta a tese da complexidade irredutível invocada pelos criacionistas, segundo a qual o maquinário celular é complexo demais para ter evoluído gradualmente. (Folha de SP, 10/4)
A pesquisa trombeteada na revista Science não falsifica a Complexidade Irredutível de Behe
A Folha de São Paulo de 10/04/2006 publicou um artigo de Kenneth Chang (New York Times) sobre um estudo publicado na revista Science que refutaria a teoria da complexidade irredutível de Behe. Em 1997 a FSP declara no seu Manual de Redação que orienta a sua conduta [sic] por um projeto editorial... com o objetivo de produzir um jornalismo crítico, moderno, pluralista e apartidário, p. 10. Nada mais falso no que diz respeito a Darwin e à Teoria do Design Inteligente (TDI), apesar das reclamações cientificamente fundamentadas enviadas aos ombudsmen desde 1998. Mais uma vez a FSP não segue o seu próprio manual de redação que recomenda ouvir o outro lado (p. 46) e nem fez o cruzamento de informações (p. 26-27). A FSP sabe como entrar em contato com os teóricos e proponentes da TDI.
Após vários anos afirmando que não há debate sobre a Teoria do Design Inteligente (TDI) três pesquisadores publicaram um artigo trazendo o debate para as páginas da última edição da revista Science. Os pesquisadores Jamie Bridgham, Sean Carroll e Joe Thornton afirmam ter demonstrado como que um sistema complexo irredutível conforme descrito por Michael Behe, pesquisador sênior do Discovery Institute, pode ter surgido como resultado de duplicação de gene e algumas mudanças de pontos mutacionais.
Isso continua na venerável tradição darwinista de fazer afirmações grandiosas baseadas em resultados insignificantes, disse o bioquímico Michael Behe, que desenvolveu a teoria da complexidade irredutível no seu livro best-seller A Caixa Preta de Darwin (Rio de Janeiro: Zahar, 1997). Não há nada no artigo que um proponente do DI pensasse estivesse além do alcance da mutação aleatória e da seleção natural. Em outras palavras, é um argumento tipo espantalho.
Se a FSP tivesse ouvido o outro lado, seus milhares de leitores tomariam conhecimento que os teóricos do DI rebateram as conclusões do artigo publicado na Science:
- How to Explain Irreducible Complexity – A Lab Manual - Fellows do CSC, 7 de abril de 2006
- The Science Stories that Fizzled (and the one that Might Have Been) - By Bruce Chapman, 7 de abril de 2006
- Irreducible Complexity Stands up to Biologist's Research Efforts - Discovery Institute, 6 de abril de 2006
- Michael Behe's response to Thornton research on irreducible complexity - ID The Future, 6 de abril de 2006
Os autores (inclusive Christoph Adami no seu comentário publicado na Science) estão definindo convenientemente a complexidade irredutível em nível abaixo, bem abaixo. Eu certamente não classificaria o sistema deles com nada próximo de complexidade irredutível (CI). Os sistemas CI que eu discuti no livro A Caixa Preta de Darwin contêm múltiplos fatores ativos de proteínas. O sistema deles, por outro lado, consiste apenas de uma proteína e a sua ligação. Embora na natureza o receptor e a ligação façam parte de um sistema maior que não tem uma função biológica, a parte daquele sistema maior que eles escolheram não faz nada sozinho. Em outras palavras, os componentes isolados que eles trabalharam não são irredutivelmente complexos.
No experimento, apenas resíduos de dois aminoácidos foram modificados, não foram adicionados novos componentes, nem componentes antigos foram removidos continuou Behe. O fato que tais resultados muito modestos são trombeteados deve-se mais, eu suspeito, à antipatia que muitos cientistas têm em relação ao DI do que ao valor intrínseco do próprio experimento.
No experimento, apenas resíduos de dois aminoácidos foram modificados, não foram adicionados novos componentes, nem componentes antigos foram removidos continuou Behe. O fato que tais resultados muito modestos são trombeteados deve-se mais, eu suspeito, à antipatia que muitos cientistas têm em relação ao DI do que ao valor intrínseco do próprio experimento.
- CSC Director Stephen C. Meyer’s initial response - 6 de abril de 2006
- Debating the Controversy that doesn’t exist - Por Paul Nelson - ID The Future, 6 de abril de 2006
O jornalismo científico da FSP não é crítico, moderno, pluralista e nem apartidário!
SCB (23/04/2006, p. 26) Fonte: http://www.scb.org.br/noticias2/exibe_noticias.asp?id=26&origem=scb_noticias
Bactérias: são complexas demais para serem ancestrais de eucariotos primitivos
Por que será que Patrick Forterre e Simonetta Gribaldo do Instituto Pasteur Institute disseram no comentário publicado no PNAS: “...we should definitely stop thinking of bacteria in terms of simple ‘lower’ organisms” [nós definitivamente devemos parar de pensar das bactérias em termos de simples organismos 'mais inferiores']?
+++++
Bacteria with a eukaryotic touch: A glimpse of ancient evolution?
Patrick Forterre a,b,1 and Simonetta Gribaldo a
aInstitut Pasteur, 25 rue du Docteur Roux, 75015 Paris, France; and bUniversité Paris Sud, Institut de Génétique et Microbiologie (IGM), Centre National de la Recherche Scientifique (CNRS), 91405, Orsay Cedex, France
The discovery in 1991 by Fuerst and Webb that the chromosome of the bacterium Gemmata obscuriglobus
(a member of Planctomycetales) is surrounded by a double membrane, mimicking a eukaryotic nucleus,
challenged the traditional prokaryote/eukaryote classification of living organisms based on cell structure (1).
Since then, such an unexpected observation has puzzled evolutionists, leading to different reactions.
...
In any case, the results of Fuerst and Devos and colleagues remind us that we should definitely stop thinking of bacteria in terms of simple “lower” organisms.
+++++
PDF gratuito do comentário de Forterre e Gribaldo no PNAS aqui.
+++++
NOTA CAUSTICANTE DESTE BLOGGER:
Posso ver os darwinistas ortodoxos, fundamentalistas, xiítas, pós-modernos, chiques e perfumados a la Dawkins, dizerem: Esta pesquisa lança luz sobre a história evolutiva dos seres vivos de uma forma nunca antes por nós imaginada, mas Darwin já tinha ideias a este respeito. Em um de seus Notebooks, Darwin previu que, blah, blah, blah, BLAH!!!
Mais e mais evidências de complexidade serão encontradas nos seres vivos. E Darwin, definitivamente (eu não pude resistir) não explica isso, mané!
+++++
Chaitin 'falou e disse': a matemática é complexidade irredutível
Gregory Chaitin considera as provas matemáticas como padrão ouro para todo o conhecimento. Ele disse em recente conferência sobre o entendimento: “Eu quero uma teorema que prove as obras da evolução deDarwin”. Ele foi mais além, e disse que está tentando desenvolver um “modelo de brinquedo” de biologia que a matemática pode representar.

A finalidade desta “metabiologia” [clique aqui e aqui] seria traduzir os princípios biológicos da evolução de um modo que permita provas tão simples, rigorosas e inevitáveis como aquelas encontradas na Física. Na verdade, Chaitin acredita que a matemática pura é complexidade irredutível, “e, portanto, mais próxima da biologia do que a física teórica”, in The Limits of Understanding.
Por causa desta declaração sobre a Biologia não ser ciência dura porque não tem leis e nem constantes como em Física, só princípios (a seleção natural) e que a matemática é complexidade irredutível [Argh, Chaitin cometeu um pecado epistêmico mortal e vai sofrer nas mãos insidiosas da KGB da Nomenklatura científica do mesmo modo que fizeram com Popper quando afirmou que a teoria da evolução através da seleção natural de Darwin não era nada mais do que metafísica] e que isso a aproxima da Biologia mais do que da Física.
Bem, Chaitin, você vai ter que se explicar para os atuais mandarins da Nomenklatura científica, pois o Michael Behe e os demais teóricos e proponentes do Design Inteligente vêm tentando dizer isso desde 1997, e [preencha o que está disponível nas declarações de organizações científicas, artigos publicações científicas que nós não podemos publicar, na internet, y otras cositas mais]...
+++++
Website de Gregory Chaitin
Pesquisas de apenas uma molécula de RNA Polimerase: mais e mais complexidade
Single-Molecule Studies of RNA Polymerase: One Singular Sensation, Every Little Step It Takes
Molecular Cell, Volume 41, Issue 3, 249-262, 4 February 2011
Copyright © 2011 Elsevier Inc. All rights reserved.
10.1016/j.molcel.2011.01.008
Authors
Matthew H. Larson,Robert Landick,Steven M. BlockSee Affiliations
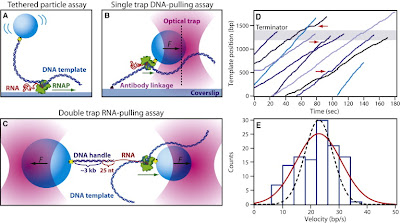
Summary
Transcription is the first of many biochemical steps that turn the genetic information found in DNA into the proteins responsible for driving cellular processes. In this review, we highlight certain advantages of single-molecule techniques in the study of prokaryotic and eukaryotic transcription, and the specific ways in which these techniques complement conventional, ensemble-based biochemistry. We focus on recent literature, highlighting examples where single-molecule methods have provided fresh insights into mechanism. We also present recent technological advances and outline future directions in the field.
+++++
FREE PDF GRATIS

Carlstadt- Administrador

- Mensagens : 1031
Idade : 48
Inscrição : 19/04/2008
Compartilhar este artigo em:
A Tese da Complexidade Irredutível :: Comentários
Qui Abr 01, 2010 10:21 pm por Carlstadt
O texto de Gabriel Cunha (UNIFESP) recomendado por Berlinck (USP) nocauteado no contexto de justificação teórica
O texto de Gabriel Cunha (UNIFESP), doutorando de Biologia Molecular, sobre os problemas da complexidade irredutível recomendado pelo Prof. Dr. Roberto G. S. Berlinck – Professor do Instituto de Química de São Carlos, da Universidade de São Paulo (USP), foi nocauteado no contexto de justificação teórica por um cruzado de esquerda do Design Inteligente
O texto de um doutorando em Biologia recomendado pelo Prof. Dr. Berlinck
Uma réplica arrasadora ao texto recomendado pelo Prof. Dr. Berlinck:
Autores de artigo do PNAS lançam mão de linguagem teleológica numa tentativa que falhou na explicação da evolução da complexidade irredutível
Sumário:
Um artigo publicado recentement no Proceedings of the National Academy of Sciences (PNAS) tenciona explicar a evolução de uma máquina molecular relativamente pequena na mitocôndria que transporte proteínas pela membrana, desse modo supostamente refutando a complexidade irredutível. Frases e afirmações como “'pre-adaptation' to bacteria ahead of a need for protein import” [pré-adaptação para a bactéria antes de uma necessidade de importar proteína], “parts accumulate until they’re ready to snap together” [as partes se acumulam até que estejam prontas para se encaixar], “machineries emerge before there’s a need for them” [as maquinarias surgem antes de haver uma necessidade para elas] ou macromutações inteligentemente “engineered” [elaboradas] são inseparáveis desta última tentativa fracassada dos críticos do Design Inteligente (DI) de refutar o argumento de complexidade irredutível de Michael Behe. Como era de se esperar quando os evolucionistas são forçados a lançar mão de linguagem e mecanismos teleológicos orientados para objetivo, isto mostra que as explicações materialistas inerentemente cegas e não guiadas não são suficientes para produzirem sistemas de complexidade irredutível. Conforme discutido mais detalhadamente abaixo, esta última tentativa de refutar a complexidade irredutível mostra inconscientemente a necessidade de design inteligene. Um resumo dos problemas inclui:
- Nenhum proponente do DI jamais afirmou que esta máquina em particular seja de complexidade irredutível, então a refutação deles é do tipo “homem de palha” [espantalho].
- A maior parte do argumento é baseada na mera evidência de homologia de sequência sem nenhuma discussão detalhada de como evoluir realmente as proteínas bacteriais em máquinas eucarióticas de transporte mitocondriais, ou resolver outros problemas necessários para evoluir esta máquina biomolecular.
- A maioria das questões sobre os passos evolucionários foi resolvida apelando-se para explicações aparentemente teleológicas que as proteínas foram magicamente “preadapted” [pré-adaptadas] para uso na máquina ou que as “machineries emerge before there’s a need for them” [as maquinarias surgem antes de haver necessidade para elas].
- Na sua única tentativa de usar uma proteína homóloga de uma bactéria no sistema eucariótico para testar a compatibilidade, os pesquisadores somente puderam fazer a função da proteína depois que eles “engineered” [elaboraram - palavra deles] inteligentemente as macromutações na proteína ao adicionarem extensas sequências de sinalização importantes bem como outras mudanças aparentemente necessárias.
- Não houve evidência nenhuma de uma versão reduzida desta máquina de transporte (“TIM”) , por isso eles tiveram que tentar encontrar uma versão reduzida de uma máquina de transporte diferente, “TOM” para fornecer “proof of principle” [prova de princípio], mas não foram mencionados os homólogos procarióticos de TOM.
- A versão supostamente mais simples de máquina de transporte TOM ainda requer múltiplas proteínas para funcionar, permitindo que ela ainda realize todas as funções essenciais, e o sistema tem pelo menos duas proteínas TOM – e não apenas uma como deu a entender o artigo do PNAS.
- Foi afirmado que o sistema TOM supostamente mais simples foi reduzido somente após uma pesquisa genômica, que os pesquisadores admitem perderia proteínas que estavam realizando funções necessárias, mas eram diferentes demais das proteínas TOM previamente conhecidas para serem detectadas.
- Conforme a filogenia padrão, o precursor supostamente mais simples nem poderia representar uma forma ancestral porque ele descendeu de um ancestral que tinha os mecanismos típicos de transporte mitocondrial.
Introdução
Artigos no SoftPedia e Wired estão trombeteando um artigo recente publicado no PNAS intitulado “The reducible complexity of a mitocondrial molecular machine” [A complexidade redutível da máquina molecular mitocondrial] afirmando que ele explica a evolução da complexidade irredutível. O parágrafo inicial do artigo do PNAS cita Michael Behe, declarando que “Proponents of Intelligent Design have argued that these sophisticated machines are 'irreducibly complex,' with this standing as the proof that, at the molecular level, Darwin’s principles of evolution cannot explain the complexity of living systems.” [Os proponentes do Design Inteligente têm argumentado que estas máquinas sofisticadas são ‘irredutivelmente complexas’, servindo isto de prova que, a nível molecular, os princípios de evolução de Darwin não podem explicar a complexidade dos sistemas vivos ]. É claro, os críticos do DI dizem constantamente que não deve existir nenhum debate científico sobre a questão da complexidade irredutível, mas não ligue pra isso.
Comecemos pelas afirmações do artigo da Softpedia sobre o flagelo bacteriano que os “basic building blocks of this ‘instrument’ can be found elsewhere in the cell as well, but performing different tasks” [os blocos básicos de construção deste ‘instrumento’ podem ser encontrados em outras partes na célula, mas realizando tarefas diferentes]. Por exemplo, todas as partes do flagelo não residem “elsewhere in the cell” [em outras partes na célula] como um número de estruturas necessárias no flagelo sem homólogos fora do flagelo. E mesmo se todas as partes residissem em algum lugar na célula, só esperando para serem cooptadas para uso num flagelo bacteriano funcional, isto resolveria muito pouco para o evolucionista. Angus Menuge explica por que estas estórias de coopção (também chamadas de “exapção”) são frágeis:
Para um flagelo funcional ser construído por exapção, as cinco condições seguintes deveriam ser todas atendidas:
C1: Disponibilidade. Entre as partes disponíveis para o recrutamento na formação do flagelo, deveriam ser aquelas capazes de realizar as tarefas altamente especializadas de pá (tipo roda propulsora de navio), rotor, e motor, muito embora todos estes itens servem alguma outra função ou nenhuma função.
C2: Sincronização. A disponibilidade destas partes teria que ser sincronizada de modo que em algum ponto, individualmente ou em combinação, todas elas estejam disponíveis ao mesmo tempo.
C3: Localização. As partes selecionadas devem todas estarem disponibilizadas ao mesmo ‘sítio de construção’, talvez não simultaneamente, mas certamente na ocasião em que forem necessitadas.
C4: Coordenação. As partes devem ser coordenadas somente no modo certo: mesmo se todas as partes de um flagelo estiverem disponíveis na ocasião certa, é claro que a maioria das maneiras de montá-las será não funcional ou irrelevante.
C5: Compatibilidade de interface. As partes devem ser mutuamente compatíveis, isto é, ‘bem casadas’ e capazes de ‘interagirem’ apropriadamente: mesmo se uma pá (tipo roda propulsora de navio), rotor, e motor forem colocadas juntas na ordem certa, elas também precisam ter interface corretamente.
(Angus Menuge, Agents Under Fire: Materialism and the Rationality of Science, p. 104-105 (Rowman & Littlefield, 2004).)
Os que se propõem explicar a evolução do flagelo quase sempre abordam somente a condição C1 e ignoram as condições C2-C5. Como será visto abaixo, esta mesma crítica se aplica com força igual aos argumentos presentes deste artigo do PNAS discutindo a origem dos mecanismos que transportam proteínas pela membrana mitocondrial.
A refutação de um homem de palha [espantalho]
As mitocondrias são um tipo de organela em células eucarióticas que produzem ATP, uma molécula que transporta eficazmente a energia utilizável para as reações celulares. Assim, as mitocondrias são frequentemente denominadas de “baterias” ou “casas de força” da célula. Elas são delimitadas por duas membranas externa e interna. Para se obter uma proteína fora da mitocondria para dentro da mitocondria, você deve passar a proteína pelas duas membranas através de vários tipos de mecanismos de transporte em cada membrana. Assim, a função destes mecanismos de transporte é pegar uma proteína e passá-la através de uma membrana. É uma função modesta, mas é uma função importante.
Basicamente, os autores do artigo do PNAS tentaram explicar como uma máquina molecular que transporta proteínas pela membrana interna da mitocondria, chamada TIM23, evoluiu. Nenhum proponente do DI jamais afirmou que este sistema particular é irredutivelmente complexo, tornando este artigo um ataque de homem de palha [espantalho]. Mas não ligue pra isso, certo?
A “Pré-adaptação” e as macromutações “elaboradas” inteligentemente resolvem as condições C2-C5?
Segundo os autores do artigo do PNAS, a evidência primária para a evolução do TIM23 é a mera semelhança entre seus componentes e outras proteínas que são encontradas em bactérias. Em um resumo-chave de seu argumento, que resolve somente a condição C1, os autores afirmam:
“We proposed that simple “core” machines were established in the first eukaryotes by drawing on preexisting bacterial proteins that had previously provided distinct functions. Subsequently, and in a step-wise process in keeping with Darwinian evolution, additional modules would have been added to the core machines to enhance their function. This proposition is supported by 3 findings: (i) that protein components found in bacteria are related in sequence to the components of mitocondrial protein transport machines, but (ii) that these bacterial proteins are not found as part of protein transport machines and (iii) that some apparently ‘‘primitive’’ organisms found today have protein transport machines that function with only one or few component parts.”
[Nós propusemos que as máquinas “centrais” simples foram estabelecidas nos primeiros eucariotos pela utilização de proteínas bacteriais pré-existentes que previamente tinham fornecido funções distintas. Subsequentemente, e em um processo gradual de acordo com a evolução darwiniana, módulos adicionais teriam sido adicionados para as máquinas centrais para melhorar sua função. Esta proposição é apoiada por 3 descobertas: (i) que os componentes encontrados nas bactérias estão relacionadas em sequência aos componentes de máquinas de transporte de proteína mitocondrial, mas (ii) que estas proteínas bacteriais não são encontradas como parte das máquinas de transporte de proteína e (iii) que alguns organismos aparentemente “primitivos” encontrados hoje têm máquinas de transporte de proteína que funcionam somente com uma ou algumas partes componentes].
Como pode ser visto, o argumento deles se baseia principalmente em encontrar homólogos de proteínas de transporte mitocondrial em algum lugar na biologia. Então, eles explicaram como que os mecanismos de transporte mitocondrial evoluíram? Dificilmente. Isto explica apenas a condição C1. Mas, para oferecer verdadeiramente “a step-wise process in keeping with Darwinian evolution” [um processo gradual de acordo com a evolução darwiniana], eles devem explicar para as condições C2-C5 e explicar como que uma vantagem funcional foi obtida ao longo de cada pequena etapa daquela evolução.
Quanto a estes homólogos, a pesquisa publicada no PNAS descobriu que 2 proteínas em TIM23 — Tim44 e Tim14 — são similares a proteínas em bactérias, TimA e TimB. TimA e TimB têm uma função diferente nas bactérias (as bactérias não têm mitocondria), mas elas residem na membrana celular bacterial, algo do tipo como a Tim44 e Tim14 residem na membrana mitocondrial interna. Foi sugerido que, se TimA e TimB se combinassem espontaneamente com uma máquina transportadora de aminoácido LivH, seria possível evoluir um complexo transportador TIM23. Eis o resumo das etapas que eles propõem:
- A evolução de uma interação binding entre TimA e LivH que permita um ponto docking para a proteína bacterial Hsp70, um motor transportador de proteína.
- Depois a evolução das interações entre TimB e LivH.
- Então, como é muito fácil fazer: “With these 3 bacterial proteins [TimA, TimB, and LivH] cooperating as subunits of a primitive transport machine, a step-wise evolution of the more sophisticated mitocondrial TIM complex would be enabled” [Com estas 3 proteínas bacteriais (TimA, TimB e LivH] cooperando como sub-unidades de uma máquina de transporte primitiva, será permitida a evolução gradual do complexo mitocondrial TIM].
O único aspecto real dos componentes explanadores requeridos de Menuge oferecido para as três proteínas é a condição C1. Eles afirmam que se as 3 partes estiverem disponíveis (etapa C1 de Menuge), então eles efetivamente resolveram o problema evolutivo. De modo algum. Quais são as probabilidades das partes se reunindo subitamente para formar um mecanismo de transporte mitocondrial funcional? Quais são as probabilidades de sítios produzindo subitamente ligação funcional entre todas estas proteínas? Estes são os principais problemas, mas eles tentaram pouco abordar estas questões. Na maior parte, eles apenas abordaram a condição C1 e declararam que o debate acabou. O argumento é algo como dizer que, se você colocar todas (ou neste caso, algumas das) partes de um computador numa caixa, e até sacudi-la um pouco, você obtém um computador. Mas as máquinas verdadeiras requerem muito mais do que apenas ter suas peças repousando numa caixa.
Há um modo de se determinar se as condições C2-C5 causam problema para a evolução destas proteínas. Eles poderiam substituir o DNA codificando Tim44 e Tim14 numa célula eucariótica de DNA com os genes codificando TimA e TimB, respectivamente, e verificar o que acontece.
Os autores não fizeram isto com TimA (eles apenas fizeram comparações sequenciais e estruturais) e, assim eles não deram realmente uma análise detalhada de como TimA pode evoluir em Tim44. Nonetheless, they assert that “Point mutations in a short segment required for interaction of the TimA protein with LivH would provide a docking point for the bacterial Hsp70, which is the direct homologue of the protein transport motor,” [Mutações pontuais em um pequeno segmento exigido para a interação da proteína TimA com LivH forneceria um ponto de ancoragem para a Hsp70 bacterial, que é o homólogo direto do motor de transporte de proteína] se bem que eles não identificam quais seriam essas mutações pontuais, ou quantas seriam necessárias existir, ou quais outras mutações (tais como a inserção de sequências de sinalização apropriadas) seriam necessárias para evoluir um proteína do tipo TimA para funcionar como Tim44.
Para TimB evoluir em Tim14, a pesquisa deles é muito mais divertida. Os autores “engineered” (elaboraram – palavra deles) uma versão modificada de TimB, pela indução de mutações, não menos importante o fato de que TimB foi “engineered for expression in yeast by adding an N-terminal mitocondrial targeting sequence and transmembrana domain” [elaborada para expressão em fermento adicionando-se uma sequência objetivando o terminal mitocondrial N e domínio de transmembrana]. Em outras palavras, eles solucionaram as condições C2, C3, e C4 – conseguindo a proteína expressada no lugar certo e no tempo certo através de algumas adições “engineered” [elaboradas] cuidadosa e inteligentemente completadas de sequências necessárias. Estes tipos de macromutações inteligentemente elaboradas não modelam o que a natureza poderia realizar razoavelmente.
Apesar disso, eles afirmam: “Although the bacterial protein function in simple assemblies, relatively little mutation would be required to convert them to function as a protein transport machine” [Embora a função da proteína bacterial em montagens simples, uma mutação relativamente pequena seria exigida para convertê-las em funcionalidade como uma máquina de transporte de proteína] A expressão “relatively little mutation” [mutação relativamente pequena] um termo técnico? Adicionar espontaneamente as sequências-alvo adequadas, os domínios de transmembrana, e as sequências de sinalização, definitivamente não é uma “relatively little mutation” [mutação relativamente pequena]. Se alguma coisa, esta pesquisa mostra que a evolução funciona melhor quando é “engineered” [elaborada] inteligentemente.
Em vez de darem uma expolicação cuidadosa das condições C2-C5, os autores do artigo do PNAS oferecem uma explicação que soa bem teleológica: “Together with the LivH amino acid transporter, these component parts would have provided ‘pre-adaptation’ to bacteria ahead of a need for protein import” [Juntas com o transportador de aminoácido LivH, estas partes componentes teriam fornecido a ‘pré-adaptação’ para as bactérias antes de uma necessidade de importação de proteínas]. Da mesma maneira, o artigo da revista Wired lida com este problema asseverando que estas partes que evoluíram, já estavam “ready to snap together” [prontas para se encaixarem] e subitamente realizaram totalmente uma nova função:
The process by which parts accumulate until they’re ready to snap together is called preadaptation. It’s a form of “neutral evolution,” in which the buildup of the parts provides no immediate advantage or disadvantage. Neutral evolution falls outside the descriptions of Charles Darwin. But once the pieces gather, mutation and natural selection can take care of the rest, ultimately resulting in the now-complex form of TIM23.[O processo pelo qual as partes se acumulam até que elas estejam prontas para se encaixarem é chamado de pré-adaptação. É uma forma de evolução neutra na qual o crescimento das partes não fornece nenhuma vantagem ou desvantagem imediats. A evolução neutra está fora das descrições de Charles Darwin. Mas, assim que as peças se reunem, a mutação e a seleção natural podem tomar conta do resto, finalmente resultando na agora complexa forma de TIM23].
A revista Wired até cita um dos cientistas que foi co-autor da pesquisa, dizendo, “But when you think about it in a neutral evolutionary fashion, in which these machineries emerge before there’s a need for them, then it makes sense” [Mas quando você pensa sobre isso de modo evolutivo neutralista, no qual estas maquinarias emergiram antes que houvesse necessidade para elas, então isto faz sentido].
Faz realmente sentido que a refutação evolucionária para a complexidade irredutível seja simplesmente pura sorte, onde, absolutamente por nenhuma razão aparente as partes desta máquina só tiveram que ser “preadapted” [pré-adaptadas] e “ready to snap together” [prontas para se encaixar] e as “machineries emerge before there’s a need for them” [maquinarias que surgem antes que haja necessidade para elas]? Se isto é a resposta para Michael Behe, então parece que os argumentos de Behe ainda estão de pé.
Expressões como “ready to snap together” [prontas para se encaixar] ou “pre-adaptation” [pré-adaptação] soa altamente como dirigida a um objetivo, e um processo cego e não-dirigido como a evolução materialista não deve ser evolução dirigida com objetivo. É claro que quando todas estas proteínas estiverem intactas e funcionando em conjunto, elas produzem uma função que dá uma vantagem. Mas, em primeiro lugar, de onde surgem subitamente estas partes, plenamente formadas, “ready to snap together” [prontas para se encaixar] para realizar esta nova função? Quando os evolucionistas invocam explicações do tipo “preadaptation” [pré-adaptação], Scott Gilbert parece estar muito certo em fazer a crítica que “[t]he modern synthesis is good at modelling the survival of the fittest, but not the arrival of the fittest” [a síntese moderna é boa em modelar a sobrevivência do mais apto, mas não a chegada do mais apto].
“Preadaptation” [pré-adpatação] ou “ready to snap together” [prontas para se encaixar] ou “machineries emerge before there’s a need for them” [maquinarias que surgem antes que haja necessidade para elas] não são explicações materialistas não-guiadas da evolução — gostem ou não, eles estão apelando inconscientemente para um processo dirigido com objetivo. E há somente um único processo dirigido com objetivo que eu conheço no pedaço: design inteligente.
Estas réplicas carregadas de teleologia me dão mais do que nunca confiança na potência do desafio da complexidade irredutível para a biologia evolutiva. São réplicas como estas que, em primeiro lugar, me fizeram cético do neodarwinismo e um proponente do Design Inteligente.
A máquina supostamente mais simples é a máquina errada e ainda exige muitas proteínas
Os autores do artigo do PNAS parecem reconhecer a improbabilidade destas partes estarem subitamente “ready to snap together” [prontas para se encaixar] e assim procuram simplificar o sistema, perguntando, “could a single component of the machine function in the absence of the others to provide even inefficient protein transport?” [poderia um só componente da função da máquina na ausência dos outros fornecer até mesmo o transporte ineficiente de proteína?] Aqui eles entram em dificuldades com respeito ao TIM23. Eles gostariam de encontrar um exemplo de apenas uma das proteínas TIM23 funcionando em algum lugar como um mecanismo de transporte pleno, mas eles não têm nenhuma evidência disso. TIM23 opera na membrana interior da mitocondria,mas existe um mecanismo de transporte similar na membrana exterior chamada TOM.
Através de uma pesquisa genômica, eles descobriram que em um caso, TOM parece ser composto de apenas 1 complexo de proteína, em vez dos costumeiros 3 ou 5 — ou como eles disseram (mais sobre isso abaixo). Isto dev ser considerado como “proof of principle” [prova do princípio] de que outras máquinas de transporte poderiam também funcionar com somente uma proteína:
Análise minuciosa da sequência do genoma de um grupo de organismos, a microsporidia, mostra que eles perderam os componentes deTom22, Tom5, Tom6,e Tom7 do seu complexo TOM central, e têm somente a sub-unidade de canal Tom40 (31).
Eles não relatarm nenhum homólogo procariótico para proteínas no complexo TOM (alguém presumiria que eles teriam relatado se tivessem encontrado-os), assim, parece que eles não encontraram nenhum exemplo de mecanismos de transporte que sejam potencialmente redutíveis e tenham homólogos procarióticos. Apesar disso, os autores make it sound como se esta função possa ser realizada com apenas uma única proteína no complexo TOM, mas levante a referência 31 deles da citação acima, e você verá uma história muito diferente e muito mais interessante.
A citação dos autores do artigo do PNAS pra este ponto, a referência 31, é um artigo de janeiro de 2009 publicado no journal Eukaryotic Cell, mostrando que o sistema de transporte mitocondrial em certas microsporidia (um filo de parasitas unicelulares) é ainda muito complexo. (Nota: as microsporidia são muito diferentes de muitos outros eucarióticos, e têm uma organela tipo mitocondria chamada de mitosoma, mas nós seguiremos a terminologia do artigo do PNAS e apenas usar o termo “mitocondria” pelos propósitos desta réplica.) Olhe a figura 5 na página 6 do artigo (gratuito [url=http://ec.asm.org/cgi/reprint/8/1/19]http://ec.asm.org/cgi/reprint/8/1/19[/url] ). No lado esquerdo na parte A está o típico sistema de transporte mitocondrial conhecido da maioria dos eucarióticos; no lado direito na parte B está o sistema para o Encephalitozoon cuniculi, a espécie de microsporidia que, supostamente, tem um mecanismo de transporte mitocondrial reduzido.
Na figura 5A, repare que o típico sistema de transporte eucariótico mitocondrial tem máquinas de transporte redundantes (i.e. 2 tipos) membrana interior, mas somente um tipo na membrana exterior. Mas no E. cuniculi, há somente um mecanismo de transporte na membrana interior. Essencialmente, esta é a extensão da “complexidade redutível” que este artigo descobriu, conforme é visto na figura 1 abaixo:
Figura 1: Reprodução altamente simplificada da figura 5 de R. F. Waller et al., “Evidence of a Reduced and Modified Mitocondrial Proteína Import Apparatus in Microsporidian Mitosomes”, Eukaryotic Cell, Vol. 8(1):19-26 (Jan. 2009). Nota: este diagrama é altamente simplificado; cada um dos complexos TIM e TOM relacionados abaixo têm um número de outras proteínas associadas com eles, bem como outros componentes necessários , no diagrama verdadeiro.

Tim23 e Tim22 têm funções e redundância que se sobrepõem para transportar proteínas através da membrana interior, como observa o artigo da Eukaryotic Cell , “Mitocondrial proteins can take one of several routes to the mitochondrion via this apparatus” [As proteínas mitocondriais podem tomar uma das diversas rotas para os mitocôndrios através deste aparato] e os complexos aparecem em “duplicate specialist form” [forma especialista duplicada]. Parece não ser absolutamente necessário ter dois tipos de máquinas de transporte na membrana interior se uma pode realizar o trabalho. Portanto, é alguma surpresa encontrar um eucarioto mais simples que tem apenas um tipo de máquina de transporte na membrana interior? Isto não é uma descoberta surpreendente pois a maioria dos eucariotos superiores parecem ter sistemas redundantes na membrana interior.
Na verdade, com respeito a algumas das proteínas supostamente não encontradas no complexo TOM do E. cuniculi na membrana exterior, o artigo destaca que “substrate overlap between Tom20 and Tom70 occurs” [substrato se sobrepõe nos complexos Tom20 e Tom70 ocorre]. Outro estudo dos complexos TIM e TOM publicado no journal Current Biology declara do mesmo modo:
“The two [outer membrana] targeting pathways are not, however, strictly separated. Some preproteins can use both Tom70–Tom37 and Tom20–Tom22 and, most importantly, preproteins that are initially recognized by Tom70 are transferred to Tom22 before their insertion into the import pore formed by Tom40 (and possibly Tom5)” [Os dois caminhos (das membranas exteriores) de targetting não são, estritamente, separados. Algumas pré-proteínas podem usar Tom70-Tom37 e Tom20-Tom22 e, mais importante, as pré-proteínas que são inicialmente reconhecidas pelo Tom70 são transferidas para Tom22 antes de sua inserção dentro do poro de importação formado por Tom40 (e possivelmente por Tom5].
Assim, há muita redundância no típico sistema de transporte eucariótico. Mas, olhando a figura 5b, no artigo, note que até no sistema “reduzido” no E. cuniculi, pelo menos 8 complexos de proteína (não uma proteína) ainda são necessários para realizarem todo o trabalho – inclusive os 4 componentes (não uma proteína) associados com a membrana exterior. E contra o artigo do PNAS, não existe apenas uma proteína no complexo TOM reduzido, mas duas.
Até onde eu sei nenhum proponente do Design Inteligente jamais afirmou que estes particulares mecanismos de transporte mitocondrial tenham complexidade irredutível. Mas, o que nós somos informados é que no E. cuniculi poderia representar um núcleo irredutível, como parece necessário para se ter uma máquina de transporte na membrana exterior e uma na membrana interior, cada uma com um número mínimo de partes para que este sistema funcione. O artigo do journal Eukaryotic Cell confirma este ponto ao afirmar:
“It is possible, however, that our observations reflect an import machinery that has been reduced in response to dramatic reductionism seen throughout microsporidian biology. It is notable that, of the subset of import machinery that the HMMs have identified, all major essential functions are represented, although only once rather than in duplicate specialist form as seen in fungi and animals (Fig. 5)”. (emphasis added) [Todavia, é possível que as nossas observações retratem uma maquinaria de importação que foi reduzida em resposta ao reducionismo dramático visto por toda a biologia microsporidiana. É notável que, da sub-série de maquinaria de importação que os HMMs identificaram, todas as principais funções essenciais estão representadas, embora somente uma vez em vez da forma especialista duplicada como é observado em fungos e animais (Fig. 5)”. (Ênfase adicionada).]
Eles estão dizendo que, embora nos eucariotos superiores exista redundância de função (“duplicate specialist form” [forma especialista duplicada]) para os sistemas de transporte, até em microsporidia, “all major essential functions are represented” [todas as principais funções essenciais estão representadas]. Assim, até com uma forma reduzida mais ainda complexa no E. cuniculi, ela é capaz de realizar todas as funções – uma grande admissão! Eles disseram mais que a configuração total mostra “evidence of a conserved system, albeit in relatively minimal form” [evidência de um sistema conservado, embora numa forma relativamente mínima]:
Nosso uso do HMM para pesquisar dados do E. cuniculi mostra que os candidatos para Tom70, Tom40, e Tim22 são identificados com grande confiança, e novos componentes adicionais do complexo TIM (Tim50 e Pam16) e o complexo SAM (Sam50) foram descobertos. Estes resultados oferecem mais evidência de um sistema de importação conservado, apesar de ser numa forma relativamente mínima (Fig. 5).
Realmente, quanto à proteína TOM70, eles afirmam, “Despite these losses, all other major structural features identified in the yeast Tom70 structure appear to have been conserved.” (emphasis added) [Apesar destas perdas, todas as principais características estruturais identificadas na estrutura Tom70 de fermento parecem ter sido conservadas (ênfase adicionada)]. Desse modo, não totalmente reduzido no sentido de que todos os principais componentes funcionais ainda estejam lá em alguma forma, e um punhado de proteínas são necessárias — na verdade, pelo menos 8 — para se obter uma proteína do exterior da mitocôndria para o interior da mitocôndria na microsporídia.
O artigo do PNAS constroi uma afirmação tipo homem de palha (espantalho) — que o sistema de transporte mitocondrial redundante em eucariotos superiores é de complexidade irredutível — e depois eles derrubam. O que nós observamos na microsporídia pode simplesmente representar um núcleo irredutível (vide aqui para uma explicação do conceito de “irreducible core” [núcleo irredutível]). Tente evoluir um sistema menos complexo do que este, e você pode não obter função nenhuma.
Na verdade, estes críticos do Design Inteligente podem ter deixado de fora do seu núcleo irredutível central um componente extremamente importante deste sistema — a sequência de sinal sobre as proteínas que serão transportadas. Assim como uma chave vira uma fechadura, um sinal sobre a proteína é tipicamente exigida para as proteínas ganharem acesso para dentro da mitocôndria através destes portais de transporte. Os autores também não consideraram a capacidade dos mecanismos darwinianos produzirem este componente fechadura-chave do sistema.
Assim, deve ser destacado que a referência 31 estuda o TOM, um sistema diferente do que aquele que o artigo do PNAS encontrou homólogos nas bactérias (TIM23). Olhando na referência 31, nós vemos o sistema de transporte mitocondrial mais simples conhecido é muito mais complexo do que o artigo do PNAS faz parecer.
Todas as proteínas TOM e TIM de E. cuniculi são atualmente detectáveis?
É importante salientar que o estudo no journal Eukaryotic Cell afirmou que o TOM no E. cuniculi foi reduzido somente após uma pesquisa genômica, que os pesquisadores admitem perderia proteínas que estavam realizando funções necessárias, mas que eram muito diferente das proteínas TOM previamente conhecidas como sendo detectadas. O E. cuniculi pode ter mais (ou até todas) as proteínas TOM padrão realizando todos os típicos papéis funcionais no sistema TOM típico, mas nós não saberíamos a partir desta pesquisa. Desta forma, o artigo do journal Eukaryotic Cell declara:
“The skeletal form of the mitosomal proteína import machinery identified here might reflect the difficulty in identifying homologues of many of the import proteínas of fungi and animals, particularly several of the small proteínas. Presently few genomic data exist for microsporidia aside from E. cuniculi, limiting the opportunity to look more broadly for mitosomal homologues in this group”. (emphasis added) [“a forma esqueletal da maquinaria de importação de proteína mitosomal identificada aqui pode retratar a dificuldade na identificação de homólogos de muitas das proteínas de importação dos fungos e animais, particularmente diversas das pequenas proteínas.
Presentemente, poucos dados genômicos existem para o microsporidia com exceção do E. cuniculi, limitando a oportunidade de se procurar mais amplamente para os homólogos mitosomais neste grupo”. (ênfase adicionada)]
Novamente, isto é uma grande admissão de que estas máquinas de transporte mitocondrial supostamente reduzidas possam muito bem ser mais complexas do que a pesquisa deles mostrou. Na verdade, o artigo observa que “many TOM and TIM proteins show weak conservation of primary sequence” [“muitas proteínas TOM e TIM mostram conservação fraca de sequência primária”]o que torna difícil identificá-las nas pesquisas genômicas. No caso de algumas das menores proteínas nos complexos TOM e TIM, o artigo debate se eles deveriam esperar encontrá-las:
“While some of the smaller proteins (e.g., the small Toms and tiny Tims) may be difficult to recognize because of the relative simplicity of these short sequences, the absence of matches for many of the import proteins is conspicuous given that they are readily recovered from other fungal and animal genomes” [“Embora algumas das menores proteínas (e.g., os pequenos Toms e os pequeníssimos Tims) possam ser difíceis de reconhecê-las por causa de sua relativa simplicidade destas sequências pequenas, a ausência de pares de muitas das proteínas de importação é conspícua considerando-se que elas são prontamente recuperadas de outros genomas de fungos e de animais”].
Em um caso eles escrevem que eles não têm certeza se a incapacidade de identificar certas proteínas nestes sistemas é devido ao fato de que elas não estão lá, ou se é porque eles são muito diferentes das proteínas equivalentes (que nós seqüenciamos e estamos usando como uma base de pesquisa) nos eucariotos superiores:
“It is unclear whether failure to identify further SAM complex proteins (Sam35, Sam37, and Mdm10) is due to poor conservation of these proteínas or to a capacity of Sam50 to act alone” [“Não está claro se o fracasso em identificar mais complexos SAM de proteínas (Sam35, Sam37, and Mdm10) ié devido à pobre conservação destas proteínas ou à capacidade de Sam50 de agir sozinho”].
Assim, mais proteínas equivalentes podem estar presentes no E. cuniculi do que foi relatado, mas elas podem não ter sido detectáveis na sua pesquisa genômica porque elas muito diferentes das proteínas de transporte atualmente conhecidas. Nós não sabemos com certeza qual é o caso porque estas pesquisas genômicas não investigam detalhadamente as atividades internas destes sistemas; eles apenas examinam as sequências de genes previamente conhecidas. Se as proteínas equivalentes estão no microsporidia, então não há razão para acreditar que ele tem um sistema de transporte mitocondrial reduzido.
O Encephalitozoon cuniculi não pode ser um precursor de eucariotos superiores
Finalmente, deve ser destacado que sob a filogenia de eucariotos prevalecente, é impossível que as máquinas de transporte de proteínas nos mitosomas do microsporidiano E. cuniculi representem algum estado antigo antes de os complexos múltiplos de TIM e TOM evoluírem em eucariotos superiores. Como o artigo no journal Eukaryotic Cell declara:
“Given that microsporidia are most closely related to fungi and therefor diverged within the fungal-animal lineage, the ancestral microsporidia most likely possessed the common Tom and Tim proteínas seen in both fungi and animals today” [“Considerando-se que os microsporidia são mais proximamente relacionados com os fungos e, portanto, divergiram dentro da linhagem fungo-animal, o microsporidia ancestral muito provavelmente possuía as proteínas Tom e Tim comuns vistas hoje tanto nos fungos como nos animais”].
Deste modo, provavelmente os mecanismos de transporte reduzidos no E. cuniculi representam perda de função do estado normal mais complicado que nós encontramos nos eucariotos mais superiores:
“The specialization of two TIM complexes early in eukaryotic evolution enabled diverse mitocondrial proteína traffic. The presence of only a single inner membrana pore in E. cuniculi suggests that microsporidia have reversed this specialization in the face of reduced proteína traffic and overall cellular reduction”(emphasis added) [“A especialização de complexos TIM no início da evolução eucariótica permitiu o tráfego mitocondrial de proteína. A presença de apenas um poro de membrana interior no E. cuniculi sugere que o microsporidia reverteu esta especialização devido ao tráfego reduzido de proteína e redução celular total”(ênfase adicionada)].
Assim, este organismo não pode na verdade mostrar diretamente alguns precursores evolucionários para os mecanismos de transporte mitocondrial mais avançados que nós observamos hoje.
Conclusão
O artigo do Wired afirma que o sistema de transporte mitocondrial “seems to pose a cellular chicken-and-egg question: How could proteína transport evolve when it was necessary to survive in the first place?” [parece propor uma questão celular do tipo quem veio primeiro o ovo ou a galinha]. Contudo, como pode ser visto, as pesquisas estão mostrando que até nos sistemas “reduzidos” de transporte mitocondrial, “all major essential functions are represented” [todas as principais funções essenciais estão representadas ] e muitas proteínas são necessárias para realizar o trabalho. A principal diferença é a falta de sistemas redundantes e muito menos total de proteínas.
O artigo do Wired não resolveu esta “questão do tipo quem veio primeiro, o ovo ou a galinha” porque até o sistema tido como mais simples ainda é complexo, contendo máquinas múltiplas funcionais de transporte de múltiplas proteínas bem como de um número de proteínas adicionais requeridas para o processo de transporte.
O que é mais revelador é que esses evolucionistas foram forçados a lançar mão de linguagem explanatória dirigida a objetivos do tipo como “preadapted” [pré-adaptado], “parts accumulate until they’re ready to snap together” [as partes se acumulam até que estejam prontas para se encaixar]ou “machineries emerge before there’s a need for them” [as maquinarias surgem antes de haver uma necessidade para elas]. No ID proponent has ever claimed these particular systems are irreducibly complex, but if these evolutionists' arguments are any indication, then even here intelligent design seems to have the upper hand, and blind and undirected processes appear insufficient. If this paper's explanation for the evolution of this machine, in their own words, “provides a blueprint for the evolution of cellular machinery in general” [fornece uma planta para a evolução da maquinaria celular em geral], estão está claro que as explicações evolutivas darwinianas para as origens da maquinaria celular são deficientes.
O artigo do PNAS afirma corretamente, “How these molecular machines evolved is a fundamental question” [Como que estas máquinas moleculares evoluíram é uma questão fundamental]. Alguém pensaria que uma questão fundamental dessas exigiria uma resposta rigorosa e detalhada. Infelizmente, os autores do artigo do PNAS tratam esta “questão fundamental” como se estivesse resolvida e a complexidade irredutível fosse refutada através de alguma investigação superficial e um punhado de explicações que parecem teleológicas. Eles querem ganhar o debate sem ter acontecido um.
Eles estão certos de que isto é uma “questão fundamental”, e a complexidade irredutível — nos casos onde os proponentes do DI têm argumentado realmente — ainda tem muito mérito.
Postado por Casey Luskin em 4 de setembro de 2009 12:36 PM | Permalink
+++++
COMENTÁRIOS IMPERTINENTES DESTE BLOGGER:
1. Berlinck, você deve recomendar aos seus leitores a leitura desta réplica. Este é ‘o outro lado da história’ do DI que nunca vemos nos blogs da rede [url=http://scienceblogs.com.br]http://scienceblogs.com.br[/url].
2. Prá finalizar, minha opinião sobre o status epistêmico da teoria de Darwin (Design não inteligente) no contexto de justificação teórica e pré-Síntese Evolutiva Ampliada (2010):

Fonte: Revista Science

Fonte: iStock Photos
3. É, minha gente, tem coisa mais bonita do que as evidências em Biologia e Química? Contrariando o mantra evolutivo de Dobzhansky, nada em biologia faz sentido, a não ser à luz das evidências.
4. Com esta longa e detalhada réplica, doravante o Prof. Dr. Berlinck (USP) deverá ler os artigos científicos e será muito mais criterioso na recomendação de artigos mencionados por terceiros que supostamente falsificam a tese de complexidade irredutível de Behe.
O texto de Gabriel Cunha (UNIFESP), doutorando de Biologia Molecular, sobre os problemas da complexidade irredutível recomendado pelo Prof. Dr. Roberto G. S. Berlinck – Professor do Instituto de Química de São Carlos, da Universidade de São Paulo (USP), foi nocauteado no contexto de justificação teórica por um cruzado de esquerda do Design Inteligente
O texto de um doutorando em Biologia recomendado pelo Prof. Dr. Berlinck
Uma réplica arrasadora ao texto recomendado pelo Prof. Dr. Berlinck:
Autores de artigo do PNAS lançam mão de linguagem teleológica numa tentativa que falhou na explicação da evolução da complexidade irredutível
Sumário:
Um artigo publicado recentement no Proceedings of the National Academy of Sciences (PNAS) tenciona explicar a evolução de uma máquina molecular relativamente pequena na mitocôndria que transporte proteínas pela membrana, desse modo supostamente refutando a complexidade irredutível. Frases e afirmações como “'pre-adaptation' to bacteria ahead of a need for protein import” [pré-adaptação para a bactéria antes de uma necessidade de importar proteína], “parts accumulate until they’re ready to snap together” [as partes se acumulam até que estejam prontas para se encaixar], “machineries emerge before there’s a need for them” [as maquinarias surgem antes de haver uma necessidade para elas] ou macromutações inteligentemente “engineered” [elaboradas] são inseparáveis desta última tentativa fracassada dos críticos do Design Inteligente (DI) de refutar o argumento de complexidade irredutível de Michael Behe. Como era de se esperar quando os evolucionistas são forçados a lançar mão de linguagem e mecanismos teleológicos orientados para objetivo, isto mostra que as explicações materialistas inerentemente cegas e não guiadas não são suficientes para produzirem sistemas de complexidade irredutível. Conforme discutido mais detalhadamente abaixo, esta última tentativa de refutar a complexidade irredutível mostra inconscientemente a necessidade de design inteligene. Um resumo dos problemas inclui:
- Nenhum proponente do DI jamais afirmou que esta máquina em particular seja de complexidade irredutível, então a refutação deles é do tipo “homem de palha” [espantalho].
- A maior parte do argumento é baseada na mera evidência de homologia de sequência sem nenhuma discussão detalhada de como evoluir realmente as proteínas bacteriais em máquinas eucarióticas de transporte mitocondriais, ou resolver outros problemas necessários para evoluir esta máquina biomolecular.
- A maioria das questões sobre os passos evolucionários foi resolvida apelando-se para explicações aparentemente teleológicas que as proteínas foram magicamente “preadapted” [pré-adaptadas] para uso na máquina ou que as “machineries emerge before there’s a need for them” [as maquinarias surgem antes de haver necessidade para elas].
- Na sua única tentativa de usar uma proteína homóloga de uma bactéria no sistema eucariótico para testar a compatibilidade, os pesquisadores somente puderam fazer a função da proteína depois que eles “engineered” [elaboraram - palavra deles] inteligentemente as macromutações na proteína ao adicionarem extensas sequências de sinalização importantes bem como outras mudanças aparentemente necessárias.
- Não houve evidência nenhuma de uma versão reduzida desta máquina de transporte (“TIM”) , por isso eles tiveram que tentar encontrar uma versão reduzida de uma máquina de transporte diferente, “TOM” para fornecer “proof of principle” [prova de princípio], mas não foram mencionados os homólogos procarióticos de TOM.
- A versão supostamente mais simples de máquina de transporte TOM ainda requer múltiplas proteínas para funcionar, permitindo que ela ainda realize todas as funções essenciais, e o sistema tem pelo menos duas proteínas TOM – e não apenas uma como deu a entender o artigo do PNAS.
- Foi afirmado que o sistema TOM supostamente mais simples foi reduzido somente após uma pesquisa genômica, que os pesquisadores admitem perderia proteínas que estavam realizando funções necessárias, mas eram diferentes demais das proteínas TOM previamente conhecidas para serem detectadas.
- Conforme a filogenia padrão, o precursor supostamente mais simples nem poderia representar uma forma ancestral porque ele descendeu de um ancestral que tinha os mecanismos típicos de transporte mitocondrial.
Introdução
Artigos no SoftPedia e Wired estão trombeteando um artigo recente publicado no PNAS intitulado “The reducible complexity of a mitocondrial molecular machine” [A complexidade redutível da máquina molecular mitocondrial] afirmando que ele explica a evolução da complexidade irredutível. O parágrafo inicial do artigo do PNAS cita Michael Behe, declarando que “Proponents of Intelligent Design have argued that these sophisticated machines are 'irreducibly complex,' with this standing as the proof that, at the molecular level, Darwin’s principles of evolution cannot explain the complexity of living systems.” [Os proponentes do Design Inteligente têm argumentado que estas máquinas sofisticadas são ‘irredutivelmente complexas’, servindo isto de prova que, a nível molecular, os princípios de evolução de Darwin não podem explicar a complexidade dos sistemas vivos ]. É claro, os críticos do DI dizem constantamente que não deve existir nenhum debate científico sobre a questão da complexidade irredutível, mas não ligue pra isso.
Comecemos pelas afirmações do artigo da Softpedia sobre o flagelo bacteriano que os “basic building blocks of this ‘instrument’ can be found elsewhere in the cell as well, but performing different tasks” [os blocos básicos de construção deste ‘instrumento’ podem ser encontrados em outras partes na célula, mas realizando tarefas diferentes]. Por exemplo, todas as partes do flagelo não residem “elsewhere in the cell” [em outras partes na célula] como um número de estruturas necessárias no flagelo sem homólogos fora do flagelo. E mesmo se todas as partes residissem em algum lugar na célula, só esperando para serem cooptadas para uso num flagelo bacteriano funcional, isto resolveria muito pouco para o evolucionista. Angus Menuge explica por que estas estórias de coopção (também chamadas de “exapção”) são frágeis:
Para um flagelo funcional ser construído por exapção, as cinco condições seguintes deveriam ser todas atendidas:
C1: Disponibilidade. Entre as partes disponíveis para o recrutamento na formação do flagelo, deveriam ser aquelas capazes de realizar as tarefas altamente especializadas de pá (tipo roda propulsora de navio), rotor, e motor, muito embora todos estes itens servem alguma outra função ou nenhuma função.
C2: Sincronização. A disponibilidade destas partes teria que ser sincronizada de modo que em algum ponto, individualmente ou em combinação, todas elas estejam disponíveis ao mesmo tempo.
C3: Localização. As partes selecionadas devem todas estarem disponibilizadas ao mesmo ‘sítio de construção’, talvez não simultaneamente, mas certamente na ocasião em que forem necessitadas.
C4: Coordenação. As partes devem ser coordenadas somente no modo certo: mesmo se todas as partes de um flagelo estiverem disponíveis na ocasião certa, é claro que a maioria das maneiras de montá-las será não funcional ou irrelevante.
C5: Compatibilidade de interface. As partes devem ser mutuamente compatíveis, isto é, ‘bem casadas’ e capazes de ‘interagirem’ apropriadamente: mesmo se uma pá (tipo roda propulsora de navio), rotor, e motor forem colocadas juntas na ordem certa, elas também precisam ter interface corretamente.
(Angus Menuge, Agents Under Fire: Materialism and the Rationality of Science, p. 104-105 (Rowman & Littlefield, 2004).)
Os que se propõem explicar a evolução do flagelo quase sempre abordam somente a condição C1 e ignoram as condições C2-C5. Como será visto abaixo, esta mesma crítica se aplica com força igual aos argumentos presentes deste artigo do PNAS discutindo a origem dos mecanismos que transportam proteínas pela membrana mitocondrial.
A refutação de um homem de palha [espantalho]
As mitocondrias são um tipo de organela em células eucarióticas que produzem ATP, uma molécula que transporta eficazmente a energia utilizável para as reações celulares. Assim, as mitocondrias são frequentemente denominadas de “baterias” ou “casas de força” da célula. Elas são delimitadas por duas membranas externa e interna. Para se obter uma proteína fora da mitocondria para dentro da mitocondria, você deve passar a proteína pelas duas membranas através de vários tipos de mecanismos de transporte em cada membrana. Assim, a função destes mecanismos de transporte é pegar uma proteína e passá-la através de uma membrana. É uma função modesta, mas é uma função importante.
Basicamente, os autores do artigo do PNAS tentaram explicar como uma máquina molecular que transporta proteínas pela membrana interna da mitocondria, chamada TIM23, evoluiu. Nenhum proponente do DI jamais afirmou que este sistema particular é irredutivelmente complexo, tornando este artigo um ataque de homem de palha [espantalho]. Mas não ligue pra isso, certo?
A “Pré-adaptação” e as macromutações “elaboradas” inteligentemente resolvem as condições C2-C5?
Segundo os autores do artigo do PNAS, a evidência primária para a evolução do TIM23 é a mera semelhança entre seus componentes e outras proteínas que são encontradas em bactérias. Em um resumo-chave de seu argumento, que resolve somente a condição C1, os autores afirmam:
“We proposed that simple “core” machines were established in the first eukaryotes by drawing on preexisting bacterial proteins that had previously provided distinct functions. Subsequently, and in a step-wise process in keeping with Darwinian evolution, additional modules would have been added to the core machines to enhance their function. This proposition is supported by 3 findings: (i) that protein components found in bacteria are related in sequence to the components of mitocondrial protein transport machines, but (ii) that these bacterial proteins are not found as part of protein transport machines and (iii) that some apparently ‘‘primitive’’ organisms found today have protein transport machines that function with only one or few component parts.”
[Nós propusemos que as máquinas “centrais” simples foram estabelecidas nos primeiros eucariotos pela utilização de proteínas bacteriais pré-existentes que previamente tinham fornecido funções distintas. Subsequentemente, e em um processo gradual de acordo com a evolução darwiniana, módulos adicionais teriam sido adicionados para as máquinas centrais para melhorar sua função. Esta proposição é apoiada por 3 descobertas: (i) que os componentes encontrados nas bactérias estão relacionadas em sequência aos componentes de máquinas de transporte de proteína mitocondrial, mas (ii) que estas proteínas bacteriais não são encontradas como parte das máquinas de transporte de proteína e (iii) que alguns organismos aparentemente “primitivos” encontrados hoje têm máquinas de transporte de proteína que funcionam somente com uma ou algumas partes componentes].
Como pode ser visto, o argumento deles se baseia principalmente em encontrar homólogos de proteínas de transporte mitocondrial em algum lugar na biologia. Então, eles explicaram como que os mecanismos de transporte mitocondrial evoluíram? Dificilmente. Isto explica apenas a condição C1. Mas, para oferecer verdadeiramente “a step-wise process in keeping with Darwinian evolution” [um processo gradual de acordo com a evolução darwiniana], eles devem explicar para as condições C2-C5 e explicar como que uma vantagem funcional foi obtida ao longo de cada pequena etapa daquela evolução.
Quanto a estes homólogos, a pesquisa publicada no PNAS descobriu que 2 proteínas em TIM23 — Tim44 e Tim14 — são similares a proteínas em bactérias, TimA e TimB. TimA e TimB têm uma função diferente nas bactérias (as bactérias não têm mitocondria), mas elas residem na membrana celular bacterial, algo do tipo como a Tim44 e Tim14 residem na membrana mitocondrial interna. Foi sugerido que, se TimA e TimB se combinassem espontaneamente com uma máquina transportadora de aminoácido LivH, seria possível evoluir um complexo transportador TIM23. Eis o resumo das etapas que eles propõem:
- A evolução de uma interação binding entre TimA e LivH que permita um ponto docking para a proteína bacterial Hsp70, um motor transportador de proteína.
- Depois a evolução das interações entre TimB e LivH.
- Então, como é muito fácil fazer: “With these 3 bacterial proteins [TimA, TimB, and LivH] cooperating as subunits of a primitive transport machine, a step-wise evolution of the more sophisticated mitocondrial TIM complex would be enabled” [Com estas 3 proteínas bacteriais (TimA, TimB e LivH] cooperando como sub-unidades de uma máquina de transporte primitiva, será permitida a evolução gradual do complexo mitocondrial TIM].
O único aspecto real dos componentes explanadores requeridos de Menuge oferecido para as três proteínas é a condição C1. Eles afirmam que se as 3 partes estiverem disponíveis (etapa C1 de Menuge), então eles efetivamente resolveram o problema evolutivo. De modo algum. Quais são as probabilidades das partes se reunindo subitamente para formar um mecanismo de transporte mitocondrial funcional? Quais são as probabilidades de sítios produzindo subitamente ligação funcional entre todas estas proteínas? Estes são os principais problemas, mas eles tentaram pouco abordar estas questões. Na maior parte, eles apenas abordaram a condição C1 e declararam que o debate acabou. O argumento é algo como dizer que, se você colocar todas (ou neste caso, algumas das) partes de um computador numa caixa, e até sacudi-la um pouco, você obtém um computador. Mas as máquinas verdadeiras requerem muito mais do que apenas ter suas peças repousando numa caixa.
Há um modo de se determinar se as condições C2-C5 causam problema para a evolução destas proteínas. Eles poderiam substituir o DNA codificando Tim44 e Tim14 numa célula eucariótica de DNA com os genes codificando TimA e TimB, respectivamente, e verificar o que acontece.
Os autores não fizeram isto com TimA (eles apenas fizeram comparações sequenciais e estruturais) e, assim eles não deram realmente uma análise detalhada de como TimA pode evoluir em Tim44. Nonetheless, they assert that “Point mutations in a short segment required for interaction of the TimA protein with LivH would provide a docking point for the bacterial Hsp70, which is the direct homologue of the protein transport motor,” [Mutações pontuais em um pequeno segmento exigido para a interação da proteína TimA com LivH forneceria um ponto de ancoragem para a Hsp70 bacterial, que é o homólogo direto do motor de transporte de proteína] se bem que eles não identificam quais seriam essas mutações pontuais, ou quantas seriam necessárias existir, ou quais outras mutações (tais como a inserção de sequências de sinalização apropriadas) seriam necessárias para evoluir um proteína do tipo TimA para funcionar como Tim44.
Para TimB evoluir em Tim14, a pesquisa deles é muito mais divertida. Os autores “engineered” (elaboraram – palavra deles) uma versão modificada de TimB, pela indução de mutações, não menos importante o fato de que TimB foi “engineered for expression in yeast by adding an N-terminal mitocondrial targeting sequence and transmembrana domain” [elaborada para expressão em fermento adicionando-se uma sequência objetivando o terminal mitocondrial N e domínio de transmembrana]. Em outras palavras, eles solucionaram as condições C2, C3, e C4 – conseguindo a proteína expressada no lugar certo e no tempo certo através de algumas adições “engineered” [elaboradas] cuidadosa e inteligentemente completadas de sequências necessárias. Estes tipos de macromutações inteligentemente elaboradas não modelam o que a natureza poderia realizar razoavelmente.
Apesar disso, eles afirmam: “Although the bacterial protein function in simple assemblies, relatively little mutation would be required to convert them to function as a protein transport machine” [Embora a função da proteína bacterial em montagens simples, uma mutação relativamente pequena seria exigida para convertê-las em funcionalidade como uma máquina de transporte de proteína] A expressão “relatively little mutation” [mutação relativamente pequena] um termo técnico? Adicionar espontaneamente as sequências-alvo adequadas, os domínios de transmembrana, e as sequências de sinalização, definitivamente não é uma “relatively little mutation” [mutação relativamente pequena]. Se alguma coisa, esta pesquisa mostra que a evolução funciona melhor quando é “engineered” [elaborada] inteligentemente.
Em vez de darem uma expolicação cuidadosa das condições C2-C5, os autores do artigo do PNAS oferecem uma explicação que soa bem teleológica: “Together with the LivH amino acid transporter, these component parts would have provided ‘pre-adaptation’ to bacteria ahead of a need for protein import” [Juntas com o transportador de aminoácido LivH, estas partes componentes teriam fornecido a ‘pré-adaptação’ para as bactérias antes de uma necessidade de importação de proteínas]. Da mesma maneira, o artigo da revista Wired lida com este problema asseverando que estas partes que evoluíram, já estavam “ready to snap together” [prontas para se encaixarem] e subitamente realizaram totalmente uma nova função:
The process by which parts accumulate until they’re ready to snap together is called preadaptation. It’s a form of “neutral evolution,” in which the buildup of the parts provides no immediate advantage or disadvantage. Neutral evolution falls outside the descriptions of Charles Darwin. But once the pieces gather, mutation and natural selection can take care of the rest, ultimately resulting in the now-complex form of TIM23.[O processo pelo qual as partes se acumulam até que elas estejam prontas para se encaixarem é chamado de pré-adaptação. É uma forma de evolução neutra na qual o crescimento das partes não fornece nenhuma vantagem ou desvantagem imediats. A evolução neutra está fora das descrições de Charles Darwin. Mas, assim que as peças se reunem, a mutação e a seleção natural podem tomar conta do resto, finalmente resultando na agora complexa forma de TIM23].
A revista Wired até cita um dos cientistas que foi co-autor da pesquisa, dizendo, “But when you think about it in a neutral evolutionary fashion, in which these machineries emerge before there’s a need for them, then it makes sense” [Mas quando você pensa sobre isso de modo evolutivo neutralista, no qual estas maquinarias emergiram antes que houvesse necessidade para elas, então isto faz sentido].
Faz realmente sentido que a refutação evolucionária para a complexidade irredutível seja simplesmente pura sorte, onde, absolutamente por nenhuma razão aparente as partes desta máquina só tiveram que ser “preadapted” [pré-adaptadas] e “ready to snap together” [prontas para se encaixar] e as “machineries emerge before there’s a need for them” [maquinarias que surgem antes que haja necessidade para elas]? Se isto é a resposta para Michael Behe, então parece que os argumentos de Behe ainda estão de pé.
Expressões como “ready to snap together” [prontas para se encaixar] ou “pre-adaptation” [pré-adaptação] soa altamente como dirigida a um objetivo, e um processo cego e não-dirigido como a evolução materialista não deve ser evolução dirigida com objetivo. É claro que quando todas estas proteínas estiverem intactas e funcionando em conjunto, elas produzem uma função que dá uma vantagem. Mas, em primeiro lugar, de onde surgem subitamente estas partes, plenamente formadas, “ready to snap together” [prontas para se encaixar] para realizar esta nova função? Quando os evolucionistas invocam explicações do tipo “preadaptation” [pré-adaptação], Scott Gilbert parece estar muito certo em fazer a crítica que “[t]he modern synthesis is good at modelling the survival of the fittest, but not the arrival of the fittest” [a síntese moderna é boa em modelar a sobrevivência do mais apto, mas não a chegada do mais apto].
“Preadaptation” [pré-adpatação] ou “ready to snap together” [prontas para se encaixar] ou “machineries emerge before there’s a need for them” [maquinarias que surgem antes que haja necessidade para elas] não são explicações materialistas não-guiadas da evolução — gostem ou não, eles estão apelando inconscientemente para um processo dirigido com objetivo. E há somente um único processo dirigido com objetivo que eu conheço no pedaço: design inteligente.
Estas réplicas carregadas de teleologia me dão mais do que nunca confiança na potência do desafio da complexidade irredutível para a biologia evolutiva. São réplicas como estas que, em primeiro lugar, me fizeram cético do neodarwinismo e um proponente do Design Inteligente.
A máquina supostamente mais simples é a máquina errada e ainda exige muitas proteínas
Os autores do artigo do PNAS parecem reconhecer a improbabilidade destas partes estarem subitamente “ready to snap together” [prontas para se encaixar] e assim procuram simplificar o sistema, perguntando, “could a single component of the machine function in the absence of the others to provide even inefficient protein transport?” [poderia um só componente da função da máquina na ausência dos outros fornecer até mesmo o transporte ineficiente de proteína?] Aqui eles entram em dificuldades com respeito ao TIM23. Eles gostariam de encontrar um exemplo de apenas uma das proteínas TIM23 funcionando em algum lugar como um mecanismo de transporte pleno, mas eles não têm nenhuma evidência disso. TIM23 opera na membrana interior da mitocondria,mas existe um mecanismo de transporte similar na membrana exterior chamada TOM.
Através de uma pesquisa genômica, eles descobriram que em um caso, TOM parece ser composto de apenas 1 complexo de proteína, em vez dos costumeiros 3 ou 5 — ou como eles disseram (mais sobre isso abaixo). Isto dev ser considerado como “proof of principle” [prova do princípio] de que outras máquinas de transporte poderiam também funcionar com somente uma proteína:
Análise minuciosa da sequência do genoma de um grupo de organismos, a microsporidia, mostra que eles perderam os componentes deTom22, Tom5, Tom6,e Tom7 do seu complexo TOM central, e têm somente a sub-unidade de canal Tom40 (31).
Eles não relatarm nenhum homólogo procariótico para proteínas no complexo TOM (alguém presumiria que eles teriam relatado se tivessem encontrado-os), assim, parece que eles não encontraram nenhum exemplo de mecanismos de transporte que sejam potencialmente redutíveis e tenham homólogos procarióticos. Apesar disso, os autores make it sound como se esta função possa ser realizada com apenas uma única proteína no complexo TOM, mas levante a referência 31 deles da citação acima, e você verá uma história muito diferente e muito mais interessante.
A citação dos autores do artigo do PNAS pra este ponto, a referência 31, é um artigo de janeiro de 2009 publicado no journal Eukaryotic Cell, mostrando que o sistema de transporte mitocondrial em certas microsporidia (um filo de parasitas unicelulares) é ainda muito complexo. (Nota: as microsporidia são muito diferentes de muitos outros eucarióticos, e têm uma organela tipo mitocondria chamada de mitosoma, mas nós seguiremos a terminologia do artigo do PNAS e apenas usar o termo “mitocondria” pelos propósitos desta réplica.) Olhe a figura 5 na página 6 do artigo (gratuito [url=http://ec.asm.org/cgi/reprint/8/1/19]http://ec.asm.org/cgi/reprint/8/1/19[/url] ). No lado esquerdo na parte A está o típico sistema de transporte mitocondrial conhecido da maioria dos eucarióticos; no lado direito na parte B está o sistema para o Encephalitozoon cuniculi, a espécie de microsporidia que, supostamente, tem um mecanismo de transporte mitocondrial reduzido.
Na figura 5A, repare que o típico sistema de transporte eucariótico mitocondrial tem máquinas de transporte redundantes (i.e. 2 tipos) membrana interior, mas somente um tipo na membrana exterior. Mas no E. cuniculi, há somente um mecanismo de transporte na membrana interior. Essencialmente, esta é a extensão da “complexidade redutível” que este artigo descobriu, conforme é visto na figura 1 abaixo:
Figura 1: Reprodução altamente simplificada da figura 5 de R. F. Waller et al., “Evidence of a Reduced and Modified Mitocondrial Proteína Import Apparatus in Microsporidian Mitosomes”, Eukaryotic Cell, Vol. 8(1):19-26 (Jan. 2009). Nota: este diagrama é altamente simplificado; cada um dos complexos TIM e TOM relacionados abaixo têm um número de outras proteínas associadas com eles, bem como outros componentes necessários , no diagrama verdadeiro.
Tim23 e Tim22 têm funções e redundância que se sobrepõem para transportar proteínas através da membrana interior, como observa o artigo da Eukaryotic Cell , “Mitocondrial proteins can take one of several routes to the mitochondrion via this apparatus” [As proteínas mitocondriais podem tomar uma das diversas rotas para os mitocôndrios através deste aparato] e os complexos aparecem em “duplicate specialist form” [forma especialista duplicada]. Parece não ser absolutamente necessário ter dois tipos de máquinas de transporte na membrana interior se uma pode realizar o trabalho. Portanto, é alguma surpresa encontrar um eucarioto mais simples que tem apenas um tipo de máquina de transporte na membrana interior? Isto não é uma descoberta surpreendente pois a maioria dos eucariotos superiores parecem ter sistemas redundantes na membrana interior.
Na verdade, com respeito a algumas das proteínas supostamente não encontradas no complexo TOM do E. cuniculi na membrana exterior, o artigo destaca que “substrate overlap between Tom20 and Tom70 occurs” [substrato se sobrepõe nos complexos Tom20 e Tom70 ocorre]. Outro estudo dos complexos TIM e TOM publicado no journal Current Biology declara do mesmo modo:
“The two [outer membrana] targeting pathways are not, however, strictly separated. Some preproteins can use both Tom70–Tom37 and Tom20–Tom22 and, most importantly, preproteins that are initially recognized by Tom70 are transferred to Tom22 before their insertion into the import pore formed by Tom40 (and possibly Tom5)” [Os dois caminhos (das membranas exteriores) de targetting não são, estritamente, separados. Algumas pré-proteínas podem usar Tom70-Tom37 e Tom20-Tom22 e, mais importante, as pré-proteínas que são inicialmente reconhecidas pelo Tom70 são transferidas para Tom22 antes de sua inserção dentro do poro de importação formado por Tom40 (e possivelmente por Tom5].
Assim, há muita redundância no típico sistema de transporte eucariótico. Mas, olhando a figura 5b, no artigo, note que até no sistema “reduzido” no E. cuniculi, pelo menos 8 complexos de proteína (não uma proteína) ainda são necessários para realizarem todo o trabalho – inclusive os 4 componentes (não uma proteína) associados com a membrana exterior. E contra o artigo do PNAS, não existe apenas uma proteína no complexo TOM reduzido, mas duas.
Até onde eu sei nenhum proponente do Design Inteligente jamais afirmou que estes particulares mecanismos de transporte mitocondrial tenham complexidade irredutível. Mas, o que nós somos informados é que no E. cuniculi poderia representar um núcleo irredutível, como parece necessário para se ter uma máquina de transporte na membrana exterior e uma na membrana interior, cada uma com um número mínimo de partes para que este sistema funcione. O artigo do journal Eukaryotic Cell confirma este ponto ao afirmar:
“It is possible, however, that our observations reflect an import machinery that has been reduced in response to dramatic reductionism seen throughout microsporidian biology. It is notable that, of the subset of import machinery that the HMMs have identified, all major essential functions are represented, although only once rather than in duplicate specialist form as seen in fungi and animals (Fig. 5)”. (emphasis added) [Todavia, é possível que as nossas observações retratem uma maquinaria de importação que foi reduzida em resposta ao reducionismo dramático visto por toda a biologia microsporidiana. É notável que, da sub-série de maquinaria de importação que os HMMs identificaram, todas as principais funções essenciais estão representadas, embora somente uma vez em vez da forma especialista duplicada como é observado em fungos e animais (Fig. 5)”. (Ênfase adicionada).]
Eles estão dizendo que, embora nos eucariotos superiores exista redundância de função (“duplicate specialist form” [forma especialista duplicada]) para os sistemas de transporte, até em microsporidia, “all major essential functions are represented” [todas as principais funções essenciais estão representadas]. Assim, até com uma forma reduzida mais ainda complexa no E. cuniculi, ela é capaz de realizar todas as funções – uma grande admissão! Eles disseram mais que a configuração total mostra “evidence of a conserved system, albeit in relatively minimal form” [evidência de um sistema conservado, embora numa forma relativamente mínima]:
Nosso uso do HMM para pesquisar dados do E. cuniculi mostra que os candidatos para Tom70, Tom40, e Tim22 são identificados com grande confiança, e novos componentes adicionais do complexo TIM (Tim50 e Pam16) e o complexo SAM (Sam50) foram descobertos. Estes resultados oferecem mais evidência de um sistema de importação conservado, apesar de ser numa forma relativamente mínima (Fig. 5).
Realmente, quanto à proteína TOM70, eles afirmam, “Despite these losses, all other major structural features identified in the yeast Tom70 structure appear to have been conserved.” (emphasis added) [Apesar destas perdas, todas as principais características estruturais identificadas na estrutura Tom70 de fermento parecem ter sido conservadas (ênfase adicionada)]. Desse modo, não totalmente reduzido no sentido de que todos os principais componentes funcionais ainda estejam lá em alguma forma, e um punhado de proteínas são necessárias — na verdade, pelo menos 8 — para se obter uma proteína do exterior da mitocôndria para o interior da mitocôndria na microsporídia.
O artigo do PNAS constroi uma afirmação tipo homem de palha (espantalho) — que o sistema de transporte mitocondrial redundante em eucariotos superiores é de complexidade irredutível — e depois eles derrubam. O que nós observamos na microsporídia pode simplesmente representar um núcleo irredutível (vide aqui para uma explicação do conceito de “irreducible core” [núcleo irredutível]). Tente evoluir um sistema menos complexo do que este, e você pode não obter função nenhuma.
Na verdade, estes críticos do Design Inteligente podem ter deixado de fora do seu núcleo irredutível central um componente extremamente importante deste sistema — a sequência de sinal sobre as proteínas que serão transportadas. Assim como uma chave vira uma fechadura, um sinal sobre a proteína é tipicamente exigida para as proteínas ganharem acesso para dentro da mitocôndria através destes portais de transporte. Os autores também não consideraram a capacidade dos mecanismos darwinianos produzirem este componente fechadura-chave do sistema.
Assim, deve ser destacado que a referência 31 estuda o TOM, um sistema diferente do que aquele que o artigo do PNAS encontrou homólogos nas bactérias (TIM23). Olhando na referência 31, nós vemos o sistema de transporte mitocondrial mais simples conhecido é muito mais complexo do que o artigo do PNAS faz parecer.
Todas as proteínas TOM e TIM de E. cuniculi são atualmente detectáveis?
É importante salientar que o estudo no journal Eukaryotic Cell afirmou que o TOM no E. cuniculi foi reduzido somente após uma pesquisa genômica, que os pesquisadores admitem perderia proteínas que estavam realizando funções necessárias, mas que eram muito diferente das proteínas TOM previamente conhecidas como sendo detectadas. O E. cuniculi pode ter mais (ou até todas) as proteínas TOM padrão realizando todos os típicos papéis funcionais no sistema TOM típico, mas nós não saberíamos a partir desta pesquisa. Desta forma, o artigo do journal Eukaryotic Cell declara:
“The skeletal form of the mitosomal proteína import machinery identified here might reflect the difficulty in identifying homologues of many of the import proteínas of fungi and animals, particularly several of the small proteínas. Presently few genomic data exist for microsporidia aside from E. cuniculi, limiting the opportunity to look more broadly for mitosomal homologues in this group”. (emphasis added) [“a forma esqueletal da maquinaria de importação de proteína mitosomal identificada aqui pode retratar a dificuldade na identificação de homólogos de muitas das proteínas de importação dos fungos e animais, particularmente diversas das pequenas proteínas.
Presentemente, poucos dados genômicos existem para o microsporidia com exceção do E. cuniculi, limitando a oportunidade de se procurar mais amplamente para os homólogos mitosomais neste grupo”. (ênfase adicionada)]
Novamente, isto é uma grande admissão de que estas máquinas de transporte mitocondrial supostamente reduzidas possam muito bem ser mais complexas do que a pesquisa deles mostrou. Na verdade, o artigo observa que “many TOM and TIM proteins show weak conservation of primary sequence” [“muitas proteínas TOM e TIM mostram conservação fraca de sequência primária”]o que torna difícil identificá-las nas pesquisas genômicas. No caso de algumas das menores proteínas nos complexos TOM e TIM, o artigo debate se eles deveriam esperar encontrá-las:
“While some of the smaller proteins (e.g., the small Toms and tiny Tims) may be difficult to recognize because of the relative simplicity of these short sequences, the absence of matches for many of the import proteins is conspicuous given that they are readily recovered from other fungal and animal genomes” [“Embora algumas das menores proteínas (e.g., os pequenos Toms e os pequeníssimos Tims) possam ser difíceis de reconhecê-las por causa de sua relativa simplicidade destas sequências pequenas, a ausência de pares de muitas das proteínas de importação é conspícua considerando-se que elas são prontamente recuperadas de outros genomas de fungos e de animais”].
Em um caso eles escrevem que eles não têm certeza se a incapacidade de identificar certas proteínas nestes sistemas é devido ao fato de que elas não estão lá, ou se é porque eles são muito diferentes das proteínas equivalentes (que nós seqüenciamos e estamos usando como uma base de pesquisa) nos eucariotos superiores:
“It is unclear whether failure to identify further SAM complex proteins (Sam35, Sam37, and Mdm10) is due to poor conservation of these proteínas or to a capacity of Sam50 to act alone” [“Não está claro se o fracasso em identificar mais complexos SAM de proteínas (Sam35, Sam37, and Mdm10) ié devido à pobre conservação destas proteínas ou à capacidade de Sam50 de agir sozinho”].
Assim, mais proteínas equivalentes podem estar presentes no E. cuniculi do que foi relatado, mas elas podem não ter sido detectáveis na sua pesquisa genômica porque elas muito diferentes das proteínas de transporte atualmente conhecidas. Nós não sabemos com certeza qual é o caso porque estas pesquisas genômicas não investigam detalhadamente as atividades internas destes sistemas; eles apenas examinam as sequências de genes previamente conhecidas. Se as proteínas equivalentes estão no microsporidia, então não há razão para acreditar que ele tem um sistema de transporte mitocondrial reduzido.
O Encephalitozoon cuniculi não pode ser um precursor de eucariotos superiores
Finalmente, deve ser destacado que sob a filogenia de eucariotos prevalecente, é impossível que as máquinas de transporte de proteínas nos mitosomas do microsporidiano E. cuniculi representem algum estado antigo antes de os complexos múltiplos de TIM e TOM evoluírem em eucariotos superiores. Como o artigo no journal Eukaryotic Cell declara:
“Given that microsporidia are most closely related to fungi and therefor diverged within the fungal-animal lineage, the ancestral microsporidia most likely possessed the common Tom and Tim proteínas seen in both fungi and animals today” [“Considerando-se que os microsporidia são mais proximamente relacionados com os fungos e, portanto, divergiram dentro da linhagem fungo-animal, o microsporidia ancestral muito provavelmente possuía as proteínas Tom e Tim comuns vistas hoje tanto nos fungos como nos animais”].
Deste modo, provavelmente os mecanismos de transporte reduzidos no E. cuniculi representam perda de função do estado normal mais complicado que nós encontramos nos eucariotos mais superiores:
“The specialization of two TIM complexes early in eukaryotic evolution enabled diverse mitocondrial proteína traffic. The presence of only a single inner membrana pore in E. cuniculi suggests that microsporidia have reversed this specialization in the face of reduced proteína traffic and overall cellular reduction”(emphasis added) [“A especialização de complexos TIM no início da evolução eucariótica permitiu o tráfego mitocondrial de proteína. A presença de apenas um poro de membrana interior no E. cuniculi sugere que o microsporidia reverteu esta especialização devido ao tráfego reduzido de proteína e redução celular total”(ênfase adicionada)].
Assim, este organismo não pode na verdade mostrar diretamente alguns precursores evolucionários para os mecanismos de transporte mitocondrial mais avançados que nós observamos hoje.
Conclusão
O artigo do Wired afirma que o sistema de transporte mitocondrial “seems to pose a cellular chicken-and-egg question: How could proteína transport evolve when it was necessary to survive in the first place?” [parece propor uma questão celular do tipo quem veio primeiro o ovo ou a galinha]. Contudo, como pode ser visto, as pesquisas estão mostrando que até nos sistemas “reduzidos” de transporte mitocondrial, “all major essential functions are represented” [todas as principais funções essenciais estão representadas ] e muitas proteínas são necessárias para realizar o trabalho. A principal diferença é a falta de sistemas redundantes e muito menos total de proteínas.
O artigo do Wired não resolveu esta “questão do tipo quem veio primeiro, o ovo ou a galinha” porque até o sistema tido como mais simples ainda é complexo, contendo máquinas múltiplas funcionais de transporte de múltiplas proteínas bem como de um número de proteínas adicionais requeridas para o processo de transporte.
O que é mais revelador é que esses evolucionistas foram forçados a lançar mão de linguagem explanatória dirigida a objetivos do tipo como “preadapted” [pré-adaptado], “parts accumulate until they’re ready to snap together” [as partes se acumulam até que estejam prontas para se encaixar]ou “machineries emerge before there’s a need for them” [as maquinarias surgem antes de haver uma necessidade para elas]. No ID proponent has ever claimed these particular systems are irreducibly complex, but if these evolutionists' arguments are any indication, then even here intelligent design seems to have the upper hand, and blind and undirected processes appear insufficient. If this paper's explanation for the evolution of this machine, in their own words, “provides a blueprint for the evolution of cellular machinery in general” [fornece uma planta para a evolução da maquinaria celular em geral], estão está claro que as explicações evolutivas darwinianas para as origens da maquinaria celular são deficientes.
O artigo do PNAS afirma corretamente, “How these molecular machines evolved is a fundamental question” [Como que estas máquinas moleculares evoluíram é uma questão fundamental]. Alguém pensaria que uma questão fundamental dessas exigiria uma resposta rigorosa e detalhada. Infelizmente, os autores do artigo do PNAS tratam esta “questão fundamental” como se estivesse resolvida e a complexidade irredutível fosse refutada através de alguma investigação superficial e um punhado de explicações que parecem teleológicas. Eles querem ganhar o debate sem ter acontecido um.
Eles estão certos de que isto é uma “questão fundamental”, e a complexidade irredutível — nos casos onde os proponentes do DI têm argumentado realmente — ainda tem muito mérito.
Postado por Casey Luskin em 4 de setembro de 2009 12:36 PM | Permalink
+++++
COMENTÁRIOS IMPERTINENTES DESTE BLOGGER:
1. Berlinck, você deve recomendar aos seus leitores a leitura desta réplica. Este é ‘o outro lado da história’ do DI que nunca vemos nos blogs da rede [url=http://scienceblogs.com.br]http://scienceblogs.com.br[/url].
2. Prá finalizar, minha opinião sobre o status epistêmico da teoria de Darwin (Design não inteligente) no contexto de justificação teórica e pré-Síntese Evolutiva Ampliada (2010):
Fonte: Revista Science
Fonte: iStock Photos
3. É, minha gente, tem coisa mais bonita do que as evidências em Biologia e Química? Contrariando o mantra evolutivo de Dobzhansky, nada em biologia faz sentido, a não ser à luz das evidências.
4. Com esta longa e detalhada réplica, doravante o Prof. Dr. Berlinck (USP) deverá ler os artigos científicos e será muito mais criterioso na recomendação de artigos mencionados por terceiros que supostamente falsificam a tese de complexidade irredutível de Behe.

Seg maio 31, 2010 4:39 pm por Eduardo
Três subsets de sequência de complexidade e sua relevância para informação biopolimérica
Segunda-feira, Maio 31, 2010
Theor Biol Med Model. 2005; 2: 29.
Published online 2005 August 11. doi: 10.1186/1742-4682-2-29.
PMCID: PMC1208958
Copyright 2005 Abel and Trevors; licensee BioMed Central Ltd.
2005 Abel and Trevors; licensee BioMed Central Ltd.
Three subsets of sequence complexity and their relevance to biopolymeric information
David L Abel1 and Jack T Trevors2
1Director, The Gene Emergence Project, The Origin-of-Life Foundation, Inc., 113 Hedgewood Dr., Greenbelt, MD 20770-1610 USA
2Professor, Department of Environmental Biology, University of Guelph, Rm 3220 Bovey Building, Guelph, Ontario, N1G 2W1, Canada
Corresponding author.
David L Abel: life@us.net; Jack T Trevors: jtrevors@uoguelph.ca
Received May 23, 2005; Accepted August 11, 2005.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Image not related to this article/Imagem não relacionada com este artigo.
California Institute for Quantitative Biosciences
Genetic algorithms instruct sophisticated biological organization. Three qualitative kinds of sequence complexity exist: random (RSC), ordered (OSC), and functional (FSC). FSC alone provides algorithmic instruction. Random and Ordered Sequence Complexities lie at opposite ends of the same bi-directional sequence complexity vector. Randomness in sequence space is defined by a lack of Kolmogorov algorithmic compressibility. A sequence is compressible because it contains redundant order and patterns. Law-like cause-and-effect determinism produces highly compressible order. Such forced ordering precludes both information retention and freedom of selection so critical to algorithmic programming and control. Functional Sequence Complexity requires this added programming dimension of uncoerced selection at successive decision nodes in the string. Shannon information theory measures the relative degrees of RSC and OSC. Shannon information theory cannot measure FSC. FSC is invariably associated with all forms of complex biofunction, including biochemical pathways, cycles, positive and negative feedback regulation, and homeostatic metabolism. The algorithmic programming of FSC, not merely its aperiodicity, accounts for biological organization. No empirical evidence exists of either RSC of OSC ever having produced a single instance of sophisticated biological organization. Organization invariably manifests FSC rather than successive random events (RSC) or low-informational self-ordering phenomena (OSC). Keywords: Self-organization, self-assembly, self-ordering, self-replication, genetic code origin, genetic information, self-catalysis.
+++++
FREE PDF GRÁTIS
+++++
NOTA DESTE BLOGGER:
Prestem muita, bastante atenção neste conceito -- Functional sequence complexity.
Ele vai dar muito o que falar em biologia evolutiva.
+++++
Vote neste blog para o prêmio TOPBLOG 2010.
Segunda-feira, Maio 31, 2010
Theor Biol Med Model. 2005; 2: 29.
Published online 2005 August 11. doi: 10.1186/1742-4682-2-29.
PMCID: PMC1208958
Copyright
2005 Abel and Trevors; licensee BioMed Central Ltd. Three subsets of sequence complexity and their relevance to biopolymeric information
David L Abel1 and Jack T Trevors2
1Director, The Gene Emergence Project, The Origin-of-Life Foundation, Inc., 113 Hedgewood Dr., Greenbelt, MD 20770-1610 USA
2Professor, Department of Environmental Biology, University of Guelph, Rm 3220 Bovey Building, Guelph, Ontario, N1G 2W1, Canada
Corresponding author.
David L Abel: life@us.net; Jack T Trevors: jtrevors@uoguelph.ca
Received May 23, 2005; Accepted August 11, 2005.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Image not related to this article/Imagem não relacionada com este artigo.
California Institute for Quantitative Biosciences
Genetic algorithms instruct sophisticated biological organization. Three qualitative kinds of sequence complexity exist: random (RSC), ordered (OSC), and functional (FSC). FSC alone provides algorithmic instruction. Random and Ordered Sequence Complexities lie at opposite ends of the same bi-directional sequence complexity vector. Randomness in sequence space is defined by a lack of Kolmogorov algorithmic compressibility. A sequence is compressible because it contains redundant order and patterns. Law-like cause-and-effect determinism produces highly compressible order. Such forced ordering precludes both information retention and freedom of selection so critical to algorithmic programming and control. Functional Sequence Complexity requires this added programming dimension of uncoerced selection at successive decision nodes in the string. Shannon information theory measures the relative degrees of RSC and OSC. Shannon information theory cannot measure FSC. FSC is invariably associated with all forms of complex biofunction, including biochemical pathways, cycles, positive and negative feedback regulation, and homeostatic metabolism. The algorithmic programming of FSC, not merely its aperiodicity, accounts for biological organization. No empirical evidence exists of either RSC of OSC ever having produced a single instance of sophisticated biological organization. Organization invariably manifests FSC rather than successive random events (RSC) or low-informational self-ordering phenomena (OSC). Keywords: Self-organization, self-assembly, self-ordering, self-replication, genetic code origin, genetic information, self-catalysis.
+++++
FREE PDF GRÁTIS
+++++
NOTA DESTE BLOGGER:
Prestem muita, bastante atenção neste conceito -- Functional sequence complexity.
Ele vai dar muito o que falar em biologia evolutiva.
+++++
Vote neste blog para o prêmio TOPBLOG 2010.
Seg Jul 05, 2010 3:43 pm por Eduardo
Mais complexidade: nanomáquinas na casa de máquinas das células
Nanomachines in the Powerhouse of the Cell: Architecture of the Largest Protein Complex of Cellular Respiration Elucidated
ScienceDaily (July 2, 2010) — Scientists of the University of Freiburg and the University of Frankfurt have elucidated the architecture of the largest protein complex of the cellular respiratory chain.They discovered an unknown mechanism of energy conversion in this molecular complex. The mechanism is required to utilize the energy contained in food.
The structural model of mitochondrial complex I provides new insights in energy conversion at nanoscale. A molecular coupling device links pump modules in the membrane arm of the huge enzyme complex.
(Credit: Image courtesy of Albert-Ludwigs-Universität Freiburg)
After ten years of research work, the x-ray crystallographic analysis of the huge and most complicated protein complex of the mitochondrial respiratory chain was successful. The complex contains more than 40 different proteins, marks the entry to cellular respiration and is thus also called mitochondrial complex I. The results are published in the current online-edition of the journal Science.
A detailed understanding of the function of complex I is of special medical interest. Dysfunction of the complex is implicated in several neurodegenerative diseases such as Parkinson´s disease or Alzheimer´s disease, and also with the physiological processes of biological aging, in general. The work of Prof. Carola Hunte of the Freiburg Institute for Biochemistry and Molecular Biology and the Freiburg excellence centre BIOSS (Centre for Biological Signalling Studies) in cooperation with Prof. Ulrich Brandt, Professor for Molecular Bioenergetics and member of the excellence centre „Macromolecular Complexes" and Dr. Volker Zickermann of his research group is a major step forward to this understanding.
The energy metabolism takes place in the so-called powerhouses of the cell, the mitochondria. They transduce the energy taken up as food into adenosine triphosphate, in short ATP, which is the universal energy currency of life. A chain of five complicated molecular machines in the mitochondrial membrane are responsible for the energy conversion. The production of ATP in mitochondria requires so many steps, as it is in principal a Knallgasreaction. In a laboratory experiment, hydrogen and oxygen gas would react in an explosion and the energy contained would be released as heat. In biological oxidation, the energy will be released by the membrane bound protein complexes of the respiratory chain in a controlled manner in small packages. Comparable to a fuel cell, this process generates an electrical membrane potential, which is the driving force of ATP synthesis. The total surface of all mitochondrial membranes in a human body covers about 14.000 square meter. This accounts for a daily production of about 65 kg of ATP.
...
Read more here/Leia mais aqui: Science Daily
+++++
Published Online July 1, 2010
Science DOI: 10.1126/science.1191046
Functional Modules and Structural Basis of Conformational Coupling in Mitochondrial Complex ICarola Hunte,1,2, 3,* Volker Zickermann,4,* Ulrich Brandt4,
Proton-pumping respiratory complex I is among the largest and most complicated membrane protein complexes. Its function is critical for efficient energy supply in aerobic cells and malfunctions are implicated in many neurodegenerative disorders. Here, we report the x-ray crystallographic analysis of mitochondrial complex I. The positions of all iron-sulfur clusters relative to the membrane arm were determined in the complete enzyme complex.The ubiquinone reduction site resides close to 30 Å above the membrane domain. The arrangement of functional modules suggests conformational coupling of redox chemistry with proton pumping and essentially excludes direct mechanisms. We suggest that a ~60 Å long helical transmission-element is critical for transducing conformational energy to proton-pumping elements in the distal module of the membrane arm.
1 Centre for Biological Signalling Studies (BIOSS), Institute for Biochemistry and Molecular Biology, University of Freiburg, Frieburg, Germany.
2 Institute of Membrane and Systems Biology, University of Leeds, Leeds, UK.
3 Department of Molecular Membrane Biology, Max Planck Institute of Biophysics, Frankfurt am Main, Germany.
4 Molecular Bioenergetics Group, Medical School, Cluster of Excellence Frankfurt "Macromolecular Complexes," Center for Membrane Proteomics, Goethe-University, Frankfurt am Main, Germany.
* These authors contributed equally to this work.
To whom correspondence should be addressed. E-mail: brandt@zbc.kgu.de
Received for publication 16 April 2010. Accepted for publication 10 June 2010.
+++++
Professores, pesquisadores e alunos de universidades públicas e privadas com acesso ao site CAPES/Periódicos podem ler gratuitamente este artigo da Science e de mais 22.440 publicações científicas.
+++++
NOTA TRIUNFANTE DESTE BLOGGER:
Cada vez mais pesquisadores encontram complexidade na natureza. Cada vez mais ele escrevem seus abstracts com linguagem teleológica. Cada vez mais corroboram as teses propostas de que sinais de inteligência são empiricamente detectados na natureza.
E ainda têm a cara de pau de dizer que a teoria do Design Inteligente é pseudociência e impede o avanço da ciência. Quem impede o avanço da ciência, caras-pálidas??? É a teoria da evolução reducionista de Darwin, e não o Design Inteligente.
Quem disse que o William Dembski não treplicou a réplica de Jeffrey Shallit e Wesley Elsberry?
Domingo, Julho 04, 2010
NOTA DESTE BLOGGER:
Está virando moda na internet no Brasil dizer que fulano, beltrano e sicrano escreveram críticas às teses da Teoria do Design e que seus teóricos enfiaram a viola no saco, e não responderam. Há razões para isso: vai ver as críticas foram realmente bem fundamentadas e a turma do TDI não teve saída (Darwin também não tem, mas aí a Nomenklatura científica e a Galera dos meninos e meninas de Darwin também enfiam suas violas no saco e ficam calados). Vai ver, as críticas nem eram bem fundamentadas que nem valia a pena respondê-las. Fico com esta última razão.
Já reportei aqui neste blog que estão servindo café requentado sobre a questão, e que não iria perder o meu tempo -- I have no time for trifling! Bem, parece que está virando também moda lá nos States. Mas como rapadura é doce, mas não é mole, a turma da TDI rebateu as críticas dos críticos. Serve também para os do Brasil. Especialmente os do blog Quiprona.
Sorry, periferia, mas está em inglês.
+++++
Intelligent Design Proponents Toil More than the Critics: A Response to Wesley Elsberry and Jeffrey Shallit
A few years back Dr. Wesley Elsberry and Dr. Jeffrey Shallit co-wrote an article, “Information Theory, Evolutionary Computation, and Dembski's ‘Complex Specified Information’,” in response to William Dembski’s 2001 book No Free Lunch: Why Specified Complexity Cannot Be Purchased without Intelligence.
No Free Lunch was something of a sequel to Dembski’s first major book, The Design Inference: Eliminating Chance through Small Probabilities (Cambridge University Press, 1998), but Dembski’s work has come a long way since that time. In this regard—and it’s not Elsberry or Shallit’s fault per se, this is just how things go—their critique is now somewhat out-dated. The computational research of Dembski and Robert Marks at the Evolutionary Informatics Lab (as well as the Biologic Institute) has preempted many lines of objection they raised. For example, Elsberry and Shallit charged that “intelligent design advocates have produced many popular books, but essentially no scientific research.” It’s doubtful that charge was accurate when they first posted their article, but no serious critic could make that charge in 2010.
Dembski has written brief replies to Shallit (see here and here) where he indicated that many of their criticisms are now outdated and that he did not see the need to write a further, detailed response. I too found many of Elsberry and Shallit’s critiques to be misguided and reflected misapplications of Dembski’s ideas. Nonetheless, I occasionally get emails from people interested in a rebuttal to their article, so I felt some written response is necessary. My response intends to be a non-exhaustive response to some of the errors in Elsberry and Shallit’s critique of Dembski. The response is titlted, "Intelligent Design Proponents Toil More than the Critics: A Response to Wesley Elsberry and Jeffrey Shallit," and it can be found here. I will highlight some of its main points over the course of 2 posts on Evolution News & Views.
Elsberry and Shallit Find False Positives by Misapplying Specified Complexity
A common theme in Elsberry and Shallit’s response to William Dembski is that they infer design prematurely before thinking carefully about whether the structure or event in question can in fact be explained by natural causes. In their article, they consistently seem unwilling to think hard about the objects being studied to sort out possible causal histories and determine what the right answer should be (i.e. was it designed or not?). Dembski would reply by saying that this is why we should study scenarios extremely carefully and not simply infer design until we have seriously explored the plausibility of natural explanations.
For example, in one example, Elsberry and Shallit cite pulsars as an example of a pattern that could give a false positive for design under Dembski’s explanatory filter. They write:
Pulsars (rapidly pulsating extraterrestrial radio sources) were discovered by Jocelyn Bell in 1967. She observed a long series of pulses of period 1.337 seconds. In at least one case the signal was tracked for 30 consecutive minutes, which would represent approximately 1340 pulses. Like the SETI sequence, this sequence was viewed as improbable (hence “complex") and specified (see Section 8), hence presumably it would constitute complex specified information and trigger a design inference. Yet spinning neutron stars, and not design, are the current explanation for pulsars.This reasoning misapplies specified complexity. Pulsars do produce regular repeating patterns, but those patterns aren’t complex. Elsberry and Shallit are simply wrong to claim that the patterns we observe from pulsars are unlikely. Consider this description of pulsar patterns from NASA:
Pulsars pulse because the rotation of the neutron star causes the radiation generated within the magnetic field to sweep in and out of our line of sight with a regular period.Thus, the “regular” patterns from pulsars are easily explained by natural law. The same holds for the “regular patterns formed by ice crystals,” which Elsberry and Shallit claim “would constitute CSI.” In Understanding Intelligent Design, Dembski writes (with Sean McDowell) why ice crystals are easily explained by natural law:
we cannot infer something was designed merely by eliminating chance. Star-shaped ice crystals, which form on cold windows, are a case in point. They form as a matter of physical necessity simply by virtue of the properties of water. An ice crystal has an ordered structure, but it does not warrant a design inference -- at least not in the same way as a Mickey Mouse landscape or Mount Rushmore. A designer may have designed the properties of water to bring about ice crystals, but such a design would be embedded in the laws of nature. The design we’re interested in is more like engineering design, which looks to particular structures rather than general processes.
(William Dembski and Sean McDowell, Understanding Intelligent Design: Everything You Need to Know in Plain Language, pp. 105-106 (Harvest House, 2008).)Likewise, the repeating pattern of atomic packing in a salt crystal is a pattern, but not a complex one. The laws of atomic packing and chemical bonding easily determine the structure of a salt crystal. In none of these cases would we infer design.
Elsberry and Shallit also claim that basalt columns could trigger a false positive for design under Dembski’s methods, because they might appear to be intelligently designed stone columns, well known from ancient ruins. They write:
Let us consider the construction of tall pillars made of hard material, such as stone columns. We wish to argue that all such pillars are due to intelligent agents. Now in every case where a pillar appears and the underlying causal story is known with the certainty Dembski demands, these pillars were constructed by intelligent agents (humans). Using Dembskian induction, we would conclude that intelligent agents must be responsible for all such pillars, including the sand pipes at Kodachrome Basin State Park in Utah and the basalt columns at the Giant's Causeway in Ireland. But this conclusion can only be retained by ruling out the circumstantial evidence in favor of accepted geological explanations for these features (ancient geysers and split volcanic flows, respectively; see [43]).But an ID theorist who properly applies ID thinking would never obtain a false positive from basalt columns. For one, it’s not true that “in every case where a [basalt column] appears and the underlying causal story is known with the certainty Dembski demands, these [columns] were constructed by intelligent agents (humans).” We can observe basalt lava flows in the present day cooing and then observe the columnar shapes caused by cooling basaltic lava. So we have a known, viable, natural explanation for these structures. Such columnar basalts are common in Eastern Washington State and I’ve observed them many times; they do NOT resemble human-designed pillars. Here are just a few reasons why basalt columns aren’t like “stone pillars” known from ancient human ruins:
First, basalt columns are generally hexagonal or pentagonal prisms and human designed pillars from ruins are generally round.
Second, basalt columns are never found standing alone but instead are found embedded in an outcrop representing a cooled lava flow. In contrast, human designed pillars from ruins can be found standing alone or resting on any type of rock, and need not be embedded in a lava flow.
Third, basalt columns are made of basalt and found near basalt lava -- the tell-tale sign. Human designed pillars can be found anywhere and are commonly made of marble or other carvable rock -- not basalt.It seems unlikely that a careful design theorist investigator would confuse basalt columns with a designed structure. The same goes for Elsberry and Shallit’s examples of rainbows, fungus “fairy rings,” and circular and polygonal cracks in rocks due to freezing. None of these entail false positives for Dembski’s methods. They are in every case easily explained via observed natural causes.
To be more precise, part of the reason why Elsberry and Shallit are uncovering alleged “false positives” is that they are constantly inferring design prematurely without carefully asking whether natural causes exist for that structure. Their examples infer design too quickly. They are not willing to do hard investigation to really determine if ID is in fact the best explanation, or if some natural explanation is superior. This flaw in their methodology will become increasingly clear as further examples are discussed. Specifically, my response to Elsberry and Shallit discusses this flaw in their reasoning with regards to Dembski’s “Nicholas Caputo” example and other examples in more detail.
Posted by Casey Luskin on June 30, 2010 5:19 PM | Permalink
+++++
NOTA IMPERTINENTE DESTE BLOGGER:
Ironia do destino??? Jeffrey Shallit foi professor de William Dembski. Shallit, muito obrigado pela grande ajuda ao Design Inteligente por ter formado um aluno de matemática tão brilhante como William Dembski! Mas este detalhe os críticos tupiniquins, geralmente, deixam de mencionar.
Shallit, quem diria, diretamente é responsável na formação acadêmica de um dos maiores teóricos do Design Inteligente.
+++++
Vote neste blog para o prêmio TOPBLOG 2010.
Nanomachines in the Powerhouse of the Cell: Architecture of the Largest Protein Complex of Cellular Respiration Elucidated
ScienceDaily (July 2, 2010) — Scientists of the University of Freiburg and the University of Frankfurt have elucidated the architecture of the largest protein complex of the cellular respiratory chain.They discovered an unknown mechanism of energy conversion in this molecular complex. The mechanism is required to utilize the energy contained in food.
The structural model of mitochondrial complex I provides new insights in energy conversion at nanoscale. A molecular coupling device links pump modules in the membrane arm of the huge enzyme complex.
(Credit: Image courtesy of Albert-Ludwigs-Universität Freiburg)
After ten years of research work, the x-ray crystallographic analysis of the huge and most complicated protein complex of the mitochondrial respiratory chain was successful. The complex contains more than 40 different proteins, marks the entry to cellular respiration and is thus also called mitochondrial complex I. The results are published in the current online-edition of the journal Science.
A detailed understanding of the function of complex I is of special medical interest. Dysfunction of the complex is implicated in several neurodegenerative diseases such as Parkinson´s disease or Alzheimer´s disease, and also with the physiological processes of biological aging, in general. The work of Prof. Carola Hunte of the Freiburg Institute for Biochemistry and Molecular Biology and the Freiburg excellence centre BIOSS (Centre for Biological Signalling Studies) in cooperation with Prof. Ulrich Brandt, Professor for Molecular Bioenergetics and member of the excellence centre „Macromolecular Complexes" and Dr. Volker Zickermann of his research group is a major step forward to this understanding.
The energy metabolism takes place in the so-called powerhouses of the cell, the mitochondria. They transduce the energy taken up as food into adenosine triphosphate, in short ATP, which is the universal energy currency of life. A chain of five complicated molecular machines in the mitochondrial membrane are responsible for the energy conversion. The production of ATP in mitochondria requires so many steps, as it is in principal a Knallgasreaction. In a laboratory experiment, hydrogen and oxygen gas would react in an explosion and the energy contained would be released as heat. In biological oxidation, the energy will be released by the membrane bound protein complexes of the respiratory chain in a controlled manner in small packages. Comparable to a fuel cell, this process generates an electrical membrane potential, which is the driving force of ATP synthesis. The total surface of all mitochondrial membranes in a human body covers about 14.000 square meter. This accounts for a daily production of about 65 kg of ATP.
...
Read more here/Leia mais aqui: Science Daily
+++++
Published Online July 1, 2010
Science DOI: 10.1126/science.1191046
Functional Modules and Structural Basis of Conformational Coupling in Mitochondrial Complex ICarola Hunte,1,2, 3,* Volker Zickermann,4,* Ulrich Brandt4,
Proton-pumping respiratory complex I is among the largest and most complicated membrane protein complexes. Its function is critical for efficient energy supply in aerobic cells and malfunctions are implicated in many neurodegenerative disorders. Here, we report the x-ray crystallographic analysis of mitochondrial complex I. The positions of all iron-sulfur clusters relative to the membrane arm were determined in the complete enzyme complex.The ubiquinone reduction site resides close to 30 Å above the membrane domain. The arrangement of functional modules suggests conformational coupling of redox chemistry with proton pumping and essentially excludes direct mechanisms. We suggest that a ~60 Å long helical transmission-element is critical for transducing conformational energy to proton-pumping elements in the distal module of the membrane arm.
1 Centre for Biological Signalling Studies (BIOSS), Institute for Biochemistry and Molecular Biology, University of Freiburg, Frieburg, Germany.
2 Institute of Membrane and Systems Biology, University of Leeds, Leeds, UK.
3 Department of Molecular Membrane Biology, Max Planck Institute of Biophysics, Frankfurt am Main, Germany.
4 Molecular Bioenergetics Group, Medical School, Cluster of Excellence Frankfurt "Macromolecular Complexes," Center for Membrane Proteomics, Goethe-University, Frankfurt am Main, Germany.
* These authors contributed equally to this work.
To whom correspondence should be addressed. E-mail: brandt@zbc.kgu.de
Received for publication 16 April 2010. Accepted for publication 10 June 2010.
+++++
Professores, pesquisadores e alunos de universidades públicas e privadas com acesso ao site CAPES/Periódicos podem ler gratuitamente este artigo da Science e de mais 22.440 publicações científicas.
+++++
NOTA TRIUNFANTE DESTE BLOGGER:
Cada vez mais pesquisadores encontram complexidade na natureza. Cada vez mais ele escrevem seus abstracts com linguagem teleológica. Cada vez mais corroboram as teses propostas de que sinais de inteligência são empiricamente detectados na natureza.
E ainda têm a cara de pau de dizer que a teoria do Design Inteligente é pseudociência e impede o avanço da ciência. Quem impede o avanço da ciência, caras-pálidas??? É a teoria da evolução reducionista de Darwin, e não o Design Inteligente.
Quem disse que o William Dembski não treplicou a réplica de Jeffrey Shallit e Wesley Elsberry?
Domingo, Julho 04, 2010
NOTA DESTE BLOGGER:
Está virando moda na internet no Brasil dizer que fulano, beltrano e sicrano escreveram críticas às teses da Teoria do Design e que seus teóricos enfiaram a viola no saco, e não responderam. Há razões para isso: vai ver as críticas foram realmente bem fundamentadas e a turma do TDI não teve saída (Darwin também não tem, mas aí a Nomenklatura científica e a Galera dos meninos e meninas de Darwin também enfiam suas violas no saco e ficam calados). Vai ver, as críticas nem eram bem fundamentadas que nem valia a pena respondê-las. Fico com esta última razão.
Já reportei aqui neste blog que estão servindo café requentado sobre a questão, e que não iria perder o meu tempo -- I have no time for trifling! Bem, parece que está virando também moda lá nos States. Mas como rapadura é doce, mas não é mole, a turma da TDI rebateu as críticas dos críticos. Serve também para os do Brasil. Especialmente os do blog Quiprona.
Sorry, periferia, mas está em inglês.
+++++
Intelligent Design Proponents Toil More than the Critics: A Response to Wesley Elsberry and Jeffrey Shallit
A few years back Dr. Wesley Elsberry and Dr. Jeffrey Shallit co-wrote an article, “Information Theory, Evolutionary Computation, and Dembski's ‘Complex Specified Information’,” in response to William Dembski’s 2001 book No Free Lunch: Why Specified Complexity Cannot Be Purchased without Intelligence.
No Free Lunch was something of a sequel to Dembski’s first major book, The Design Inference: Eliminating Chance through Small Probabilities (Cambridge University Press, 1998), but Dembski’s work has come a long way since that time. In this regard—and it’s not Elsberry or Shallit’s fault per se, this is just how things go—their critique is now somewhat out-dated. The computational research of Dembski and Robert Marks at the Evolutionary Informatics Lab (as well as the Biologic Institute) has preempted many lines of objection they raised. For example, Elsberry and Shallit charged that “intelligent design advocates have produced many popular books, but essentially no scientific research.” It’s doubtful that charge was accurate when they first posted their article, but no serious critic could make that charge in 2010.
Dembski has written brief replies to Shallit (see here and here) where he indicated that many of their criticisms are now outdated and that he did not see the need to write a further, detailed response. I too found many of Elsberry and Shallit’s critiques to be misguided and reflected misapplications of Dembski’s ideas. Nonetheless, I occasionally get emails from people interested in a rebuttal to their article, so I felt some written response is necessary. My response intends to be a non-exhaustive response to some of the errors in Elsberry and Shallit’s critique of Dembski. The response is titlted, "Intelligent Design Proponents Toil More than the Critics: A Response to Wesley Elsberry and Jeffrey Shallit," and it can be found here. I will highlight some of its main points over the course of 2 posts on Evolution News & Views.
Elsberry and Shallit Find False Positives by Misapplying Specified Complexity
A common theme in Elsberry and Shallit’s response to William Dembski is that they infer design prematurely before thinking carefully about whether the structure or event in question can in fact be explained by natural causes. In their article, they consistently seem unwilling to think hard about the objects being studied to sort out possible causal histories and determine what the right answer should be (i.e. was it designed or not?). Dembski would reply by saying that this is why we should study scenarios extremely carefully and not simply infer design until we have seriously explored the plausibility of natural explanations.
For example, in one example, Elsberry and Shallit cite pulsars as an example of a pattern that could give a false positive for design under Dembski’s explanatory filter. They write:
Pulsars (rapidly pulsating extraterrestrial radio sources) were discovered by Jocelyn Bell in 1967. She observed a long series of pulses of period 1.337 seconds. In at least one case the signal was tracked for 30 consecutive minutes, which would represent approximately 1340 pulses. Like the SETI sequence, this sequence was viewed as improbable (hence “complex") and specified (see Section 8), hence presumably it would constitute complex specified information and trigger a design inference. Yet spinning neutron stars, and not design, are the current explanation for pulsars.This reasoning misapplies specified complexity. Pulsars do produce regular repeating patterns, but those patterns aren’t complex. Elsberry and Shallit are simply wrong to claim that the patterns we observe from pulsars are unlikely. Consider this description of pulsar patterns from NASA:
Pulsars pulse because the rotation of the neutron star causes the radiation generated within the magnetic field to sweep in and out of our line of sight with a regular period.Thus, the “regular” patterns from pulsars are easily explained by natural law. The same holds for the “regular patterns formed by ice crystals,” which Elsberry and Shallit claim “would constitute CSI.” In Understanding Intelligent Design, Dembski writes (with Sean McDowell) why ice crystals are easily explained by natural law:
we cannot infer something was designed merely by eliminating chance. Star-shaped ice crystals, which form on cold windows, are a case in point. They form as a matter of physical necessity simply by virtue of the properties of water. An ice crystal has an ordered structure, but it does not warrant a design inference -- at least not in the same way as a Mickey Mouse landscape or Mount Rushmore. A designer may have designed the properties of water to bring about ice crystals, but such a design would be embedded in the laws of nature. The design we’re interested in is more like engineering design, which looks to particular structures rather than general processes.
(William Dembski and Sean McDowell, Understanding Intelligent Design: Everything You Need to Know in Plain Language, pp. 105-106 (Harvest House, 2008).)Likewise, the repeating pattern of atomic packing in a salt crystal is a pattern, but not a complex one. The laws of atomic packing and chemical bonding easily determine the structure of a salt crystal. In none of these cases would we infer design.
Elsberry and Shallit also claim that basalt columns could trigger a false positive for design under Dembski’s methods, because they might appear to be intelligently designed stone columns, well known from ancient ruins. They write:
Let us consider the construction of tall pillars made of hard material, such as stone columns. We wish to argue that all such pillars are due to intelligent agents. Now in every case where a pillar appears and the underlying causal story is known with the certainty Dembski demands, these pillars were constructed by intelligent agents (humans). Using Dembskian induction, we would conclude that intelligent agents must be responsible for all such pillars, including the sand pipes at Kodachrome Basin State Park in Utah and the basalt columns at the Giant's Causeway in Ireland. But this conclusion can only be retained by ruling out the circumstantial evidence in favor of accepted geological explanations for these features (ancient geysers and split volcanic flows, respectively; see [43]).But an ID theorist who properly applies ID thinking would never obtain a false positive from basalt columns. For one, it’s not true that “in every case where a [basalt column] appears and the underlying causal story is known with the certainty Dembski demands, these [columns] were constructed by intelligent agents (humans).” We can observe basalt lava flows in the present day cooing and then observe the columnar shapes caused by cooling basaltic lava. So we have a known, viable, natural explanation for these structures. Such columnar basalts are common in Eastern Washington State and I’ve observed them many times; they do NOT resemble human-designed pillars. Here are just a few reasons why basalt columns aren’t like “stone pillars” known from ancient human ruins:
First, basalt columns are generally hexagonal or pentagonal prisms and human designed pillars from ruins are generally round.
Second, basalt columns are never found standing alone but instead are found embedded in an outcrop representing a cooled lava flow. In contrast, human designed pillars from ruins can be found standing alone or resting on any type of rock, and need not be embedded in a lava flow.
Third, basalt columns are made of basalt and found near basalt lava -- the tell-tale sign. Human designed pillars can be found anywhere and are commonly made of marble or other carvable rock -- not basalt.It seems unlikely that a careful design theorist investigator would confuse basalt columns with a designed structure. The same goes for Elsberry and Shallit’s examples of rainbows, fungus “fairy rings,” and circular and polygonal cracks in rocks due to freezing. None of these entail false positives for Dembski’s methods. They are in every case easily explained via observed natural causes.
To be more precise, part of the reason why Elsberry and Shallit are uncovering alleged “false positives” is that they are constantly inferring design prematurely without carefully asking whether natural causes exist for that structure. Their examples infer design too quickly. They are not willing to do hard investigation to really determine if ID is in fact the best explanation, or if some natural explanation is superior. This flaw in their methodology will become increasingly clear as further examples are discussed. Specifically, my response to Elsberry and Shallit discusses this flaw in their reasoning with regards to Dembski’s “Nicholas Caputo” example and other examples in more detail.
Posted by Casey Luskin on June 30, 2010 5:19 PM | Permalink
+++++
NOTA IMPERTINENTE DESTE BLOGGER:
Ironia do destino??? Jeffrey Shallit foi professor de William Dembski. Shallit, muito obrigado pela grande ajuda ao Design Inteligente por ter formado um aluno de matemática tão brilhante como William Dembski! Mas este detalhe os críticos tupiniquins, geralmente, deixam de mencionar.
Shallit, quem diria, diretamente é responsável na formação acadêmica de um dos maiores teóricos do Design Inteligente.
+++++
Vote neste blog para o prêmio TOPBLOG 2010.
Ter Jul 06, 2010 10:08 am por Eduardo
Mais maquinaria extremamente complexa nos complexos de proteínas das membranas
Terça-feira, Julho 06, 2010
Published Online July 1, 2010
Science DOI: 10.1126/science.1191046
Functional Modules and Structural Basis of Conformational Coupling in Mitochondrial Complex I
Carola Hunte,1,2, 3,* Volker Zickermann,4,* Ulrich Brandt4,
Image not related to this article/Imagem não relacionada com este artigo
Proton-pumping respiratory complex I is among the largest and most complicated membrane protein complexes. Its function is critical for efficient energy supply in aerobic cells and malfunctions are implicated in many neurodegenerative disorders. Here, we report the x-ray crystallographic analysis of mitochondrial complex I. The positions of all iron-sulfur clusters relative to the membrane arm were determined in the complete enzyme complex.The ubiquinone reduction site resides close to 30 Å above the membrane domain. The arrangement of functional modules suggests conformational coupling of redox chemistry with proton pumping and essentially excludes direct mechanisms. We suggest that a ~60 Å long helical transmission-element is critical for transducing conformational energy to proton-pumping elements in the distal module of the membrane arm.
1 Centre for Biological Signalling Studies (BIOSS), Institute for Biochemistry and Molecular Biology, University of Freiburg, Frieburg, Germany.
2 Institute of Membrane and Systems Biology, University of Leeds, Leeds, UK.
3 Department of Molecular Membrane Biology, Max Planck Institute of Biophysics, Frankfurt am Main, Germany.
4 Molecular Bioenergetics Group, Medical School, Cluster of Excellence Frankfurt "Macromolecular Complexes," Center for Membrane Proteomics, Goethe-University, Frankfurt am Main, Germany.
* These authors contributed equally to this work.
To whom correspondence should be addressed. E-mail: brandt@zbc.kgu.de
+++++
Professores, pesquisadores e alunos de universidades públicas e privadas com acesso ao site CAPES/Periódicos podem ler gratuitamente este artigo da Science e de mais 22.440 publicações científicas.
+++++
PERGUNTA IMPERTINENTE DESTE BLOGGER:
Dizem que uma imagem vale mais do que mil palavras. A imagem nos remete a que na natureza? Mero acaso? Fortuita necessidade? ou Design Inteligente??? Por muito tempo eu fiquei mesmerizado pela explicação de Darwin (o homem que teve a maior ideia que toda a humanidade já teve) da evolução através da seleção natural. Todavia, depois que li o livro A Caixa Preta de Darwin, de Michael Behe, postulando que o gradualismo darwiniano não explica a origem de um 'simples' flagelo bacteriano, vi a fragilidade epistêmica de Darwin num contexto de justificação teórica que eu nunca tinha ousado perguntar.
Fui, nem sei por que, sabendo que a ciência é a busca pela verdade e que os cientistas devem seguir as evidências aonde elas forem dar. As pesquisas como esta estão demonstrando que as evidências não indo em direção a Down, mas nas direção ao Design Inteligente.
+++++
Vote neste blog para o prêmio TOPBLOG 2010.
Integrando a complexidade: o ambiente e a história - Workshop ISHPSSB 2010 - Ontario, Canadá

About the Conference
JOIN US for an interdisciplinary conference from October 7-10, at the University of Western Ontario in London, Ontario, Canada. Integrating Complexity: Environment and History will consist of two linked workshops exploring a set of challenges to scientific understanding that span many fields of the natural and human sciences, and that have broad implications for research choices, for social policy, and for how we understand ourselves and the world.
Many scientists and philosophers have treated the discovery of universal laws of nature as the primary goal of scientific research. This is often achieved via the use of experimental methods that allow scientists to study variation in particular properties of systems or to isolate particularly revealing phenomena, and the use of simplified abstract models. However, this goal has long been recognized as problematic in many areas of investigation, such as those that treat complex, experimentally inaccessible, or historically unique phenomena.
Two linked workshops will take up related themes concerning scientific understanding of complex phenomena: 1) the conceptual integration of complexity in explorations of organism-environment interaction in the life and social sciences, and 2) the methodological integration of complexity in the distinctive approaches to confirmation and explanation in the historical sciences.
The workshops will engage these issues from a variety of disciplinary perspectives, bringing together leading researchers, young scholars, and graduate students from philosophy, history, and the sciences. This diverse group of scholars will contribute to developing a new account of scientific practice that recognizes the distinctive conceptual and methodological challenges of integrating complexity. We hope that each group will gain by learning how similar conceptual and methodological problems have been handled in different fields, and how responses to them have developed historically.
The conference will be held at Western's Spencer-Ivey Centre, a venue with outstanding meeting facilities, award-winning cuisine, comfortable guest rooms, and wonderful settings for informal discussion both indoors and out.
+++++
Call for Abstracts:
The 4-day event comprises two linked workshops addressing interrelated strands in approaches to phenomena whose causal complexity poses special challenges to scientific understanding:
(1) the conceptual integration of complexity in explorations of organism-environment interaction in the life sciences and social sciences, and
(2) the methodological integration of complexity in the distinctive approaches to confirmation and explanation in the historical sciences.
The two workshops will draw together philosophers, historians, and natural and social scientists whose work focuses on the interpretation of the organism-environment interaction and its implications for our understanding of complex biological and social systems, and on the distinctive methodological challenges posed by the historical sciences.
1. Organism-Environment Interaction: Past, Present & Future
The partitioning of the complexity of the world into organism and environment—a step toward simpler abstract representations of organic phenomena—opened a space for exploration of the complex and dynamic relationship between the two. Conceptions of this relationship have played a crucial role in the development of many areas of biology, including evolutionary theory, ecology and ethology. Other disciplines, including the social sciences and medicine, have also made use of the diverse representations of the complex interaction between organisms and their environment. These relationships remain deeply contested in these domains today.
Possible topics include, but are not limited to:
The distinctive epistemological challenges of the "historical sciences" give rise to methodological debates concerning contingency, historicity, irreversibility and the limits of simple equilibrium models in biology but also in other disciplines such as palaeontology, geology, cosmology and history itself.. The implications of these issues are far-reaching for questions of evidence, explanation and the nature of understanding in the historical sciences.
Possible Topics include but are not limited to:
The event will combine plenary sessions presented by invited speakers, panel sessions, and discussion sessions led by graduate students and junior scholars.
Submission Guidelines:
Please submit a 1-page abstract for a presentation of no more than 20 minutes. There are two types of sessions: break-out discussions and panels. In a break-out discussion, the presentation introduces material for an extended discussion led by the presenter. In a panel, three related presentations precede a combined discussion. You may specify which format you prefer, though we will consider all submissions for both formats. For break-out discussions, please provide references for 2-3 background readings that can be made available in advance. This event aims to foster lively discussion across disciplinary lines and between junior and senior scholars - submissions from any relevant field and from graduate students are welcome.
Please send submissions or any inquiries to: iceh@uwo.ca
+++++
NOTA DESTE BLOGGER:
Muitos darwinistas ficam estupefatos, inclusive o Dr. Roberto Berlinck, quando eu afirmo neste blog que a teoria da evolução através da seleção natural, assim como a teoria do Design Inteligente, é uma teoria de longo alcance histórico -- tenta reconstruir eventos únicos e não repetíveis.
Todavia, de tanto ouvirem a retórica do mantra dobzhanskyano de que nada em biologia faz sentido a não à luz de evolução, eles nem são capazes de perceber que a biologia evolutiva não faz parte das ciências duras como a física que tem leis que podem ser mensuradas e suas experiências repetidas universalmente. A biologia evolutiva não tem leis, tem apenas um princípio: o da seleção natural.
QED: Toda e qualquer crítica à TDI é, diretamente, uma crítica à teoria da evolução qua ciência histórica.
A teoria do Design Inteligente propõe que a complexidade é um sinal de inteligência empiricamente detectado na natureza. E agora o pessoal do ISHPSSB vai abordar a complexidade? Bons ventos epistêmicos começam a soprar na Academia. Quando vai soprar sobre a Nomenklatura científica tupiniquim???
Será que este evento vai receber destaque posterior no boletim da ABFHIB? Duvido, mas é capaz que eu tenha de morder a minha língua...
+++++
Vote neste blog para o prêmio TOPBLOG 2010.
Terça-feira, Julho 06, 2010
Published Online July 1, 2010
Science DOI: 10.1126/science.1191046
Functional Modules and Structural Basis of Conformational Coupling in Mitochondrial Complex I
Carola Hunte,1,2, 3,* Volker Zickermann,4,* Ulrich Brandt4,
Image not related to this article/Imagem não relacionada com este artigo
Proton-pumping respiratory complex I is among the largest and most complicated membrane protein complexes. Its function is critical for efficient energy supply in aerobic cells and malfunctions are implicated in many neurodegenerative disorders. Here, we report the x-ray crystallographic analysis of mitochondrial complex I. The positions of all iron-sulfur clusters relative to the membrane arm were determined in the complete enzyme complex.The ubiquinone reduction site resides close to 30 Å above the membrane domain. The arrangement of functional modules suggests conformational coupling of redox chemistry with proton pumping and essentially excludes direct mechanisms. We suggest that a ~60 Å long helical transmission-element is critical for transducing conformational energy to proton-pumping elements in the distal module of the membrane arm.
1 Centre for Biological Signalling Studies (BIOSS), Institute for Biochemistry and Molecular Biology, University of Freiburg, Frieburg, Germany.
2 Institute of Membrane and Systems Biology, University of Leeds, Leeds, UK.
3 Department of Molecular Membrane Biology, Max Planck Institute of Biophysics, Frankfurt am Main, Germany.
4 Molecular Bioenergetics Group, Medical School, Cluster of Excellence Frankfurt "Macromolecular Complexes," Center for Membrane Proteomics, Goethe-University, Frankfurt am Main, Germany.
* These authors contributed equally to this work.
To whom correspondence should be addressed. E-mail: brandt@zbc.kgu.de
+++++
Professores, pesquisadores e alunos de universidades públicas e privadas com acesso ao site CAPES/Periódicos podem ler gratuitamente este artigo da Science e de mais 22.440 publicações científicas.
+++++
PERGUNTA IMPERTINENTE DESTE BLOGGER:
Dizem que uma imagem vale mais do que mil palavras. A imagem nos remete a que na natureza? Mero acaso? Fortuita necessidade? ou Design Inteligente??? Por muito tempo eu fiquei mesmerizado pela explicação de Darwin (o homem que teve a maior ideia que toda a humanidade já teve) da evolução através da seleção natural. Todavia, depois que li o livro A Caixa Preta de Darwin, de Michael Behe, postulando que o gradualismo darwiniano não explica a origem de um 'simples' flagelo bacteriano, vi a fragilidade epistêmica de Darwin num contexto de justificação teórica que eu nunca tinha ousado perguntar.
Fui, nem sei por que, sabendo que a ciência é a busca pela verdade e que os cientistas devem seguir as evidências aonde elas forem dar. As pesquisas como esta estão demonstrando que as evidências não indo em direção a Down, mas nas direção ao Design Inteligente.
+++++
Vote neste blog para o prêmio TOPBLOG 2010.
Integrando a complexidade: o ambiente e a história - Workshop ISHPSSB 2010 - Ontario, Canadá
About the Conference
JOIN US for an interdisciplinary conference from October 7-10, at the University of Western Ontario in London, Ontario, Canada. Integrating Complexity: Environment and History will consist of two linked workshops exploring a set of challenges to scientific understanding that span many fields of the natural and human sciences, and that have broad implications for research choices, for social policy, and for how we understand ourselves and the world.
Many scientists and philosophers have treated the discovery of universal laws of nature as the primary goal of scientific research. This is often achieved via the use of experimental methods that allow scientists to study variation in particular properties of systems or to isolate particularly revealing phenomena, and the use of simplified abstract models. However, this goal has long been recognized as problematic in many areas of investigation, such as those that treat complex, experimentally inaccessible, or historically unique phenomena.
Two linked workshops will take up related themes concerning scientific understanding of complex phenomena: 1) the conceptual integration of complexity in explorations of organism-environment interaction in the life and social sciences, and 2) the methodological integration of complexity in the distinctive approaches to confirmation and explanation in the historical sciences.
The workshops will engage these issues from a variety of disciplinary perspectives, bringing together leading researchers, young scholars, and graduate students from philosophy, history, and the sciences. This diverse group of scholars will contribute to developing a new account of scientific practice that recognizes the distinctive conceptual and methodological challenges of integrating complexity. We hope that each group will gain by learning how similar conceptual and methodological problems have been handled in different fields, and how responses to them have developed historically.
The conference will be held at Western's Spencer-Ivey Centre, a venue with outstanding meeting facilities, award-winning cuisine, comfortable guest rooms, and wonderful settings for informal discussion both indoors and out.
+++++
Call for Abstracts:
The 4-day event comprises two linked workshops addressing interrelated strands in approaches to phenomena whose causal complexity poses special challenges to scientific understanding:
(1) the conceptual integration of complexity in explorations of organism-environment interaction in the life sciences and social sciences, and
(2) the methodological integration of complexity in the distinctive approaches to confirmation and explanation in the historical sciences.
The two workshops will draw together philosophers, historians, and natural and social scientists whose work focuses on the interpretation of the organism-environment interaction and its implications for our understanding of complex biological and social systems, and on the distinctive methodological challenges posed by the historical sciences.
1. Organism-Environment Interaction: Past, Present & Future
The partitioning of the complexity of the world into organism and environment—a step toward simpler abstract representations of organic phenomena—opened a space for exploration of the complex and dynamic relationship between the two. Conceptions of this relationship have played a crucial role in the development of many areas of biology, including evolutionary theory, ecology and ethology. Other disciplines, including the social sciences and medicine, have also made use of the diverse representations of the complex interaction between organisms and their environment. These relationships remain deeply contested in these domains today.
Possible topics include, but are not limited to:
- Pre-Darwinian Conceptions of O-E Interaction
- Organization, Regulation & Response in O-E Interaction
- O-E Interaction in Evolution
- Intervening in the Environment
- O-E Interaction in Medicine
- History Matters: O-E Interaction and Historicity
- O-E Interaction in the Human Sciences
The distinctive epistemological challenges of the "historical sciences" give rise to methodological debates concerning contingency, historicity, irreversibility and the limits of simple equilibrium models in biology but also in other disciplines such as palaeontology, geology, cosmology and history itself.. The implications of these issues are far-reaching for questions of evidence, explanation and the nature of understanding in the historical sciences.
Possible Topics include but are not limited to:
- Path dependence, contingency and irreversibility
- Historical explanation
- Anthropic principles
- Reconstructing the past
- Understanding unique events
The event will combine plenary sessions presented by invited speakers, panel sessions, and discussion sessions led by graduate students and junior scholars.
Submission Guidelines:
Please submit a 1-page abstract for a presentation of no more than 20 minutes. There are two types of sessions: break-out discussions and panels. In a break-out discussion, the presentation introduces material for an extended discussion led by the presenter. In a panel, three related presentations precede a combined discussion. You may specify which format you prefer, though we will consider all submissions for both formats. For break-out discussions, please provide references for 2-3 background readings that can be made available in advance. This event aims to foster lively discussion across disciplinary lines and between junior and senior scholars - submissions from any relevant field and from graduate students are welcome.
Please send submissions or any inquiries to: iceh@uwo.ca
+++++
NOTA DESTE BLOGGER:
Muitos darwinistas ficam estupefatos, inclusive o Dr. Roberto Berlinck, quando eu afirmo neste blog que a teoria da evolução através da seleção natural, assim como a teoria do Design Inteligente, é uma teoria de longo alcance histórico -- tenta reconstruir eventos únicos e não repetíveis.
Todavia, de tanto ouvirem a retórica do mantra dobzhanskyano de que nada em biologia faz sentido a não à luz de evolução, eles nem são capazes de perceber que a biologia evolutiva não faz parte das ciências duras como a física que tem leis que podem ser mensuradas e suas experiências repetidas universalmente. A biologia evolutiva não tem leis, tem apenas um princípio: o da seleção natural.
QED: Toda e qualquer crítica à TDI é, diretamente, uma crítica à teoria da evolução qua ciência histórica.
A teoria do Design Inteligente propõe que a complexidade é um sinal de inteligência empiricamente detectado na natureza. E agora o pessoal do ISHPSSB vai abordar a complexidade? Bons ventos epistêmicos começam a soprar na Academia. Quando vai soprar sobre a Nomenklatura científica tupiniquim???
Será que este evento vai receber destaque posterior no boletim da ABFHIB? Duvido, mas é capaz que eu tenha de morder a minha língua...
+++++
Vote neste blog para o prêmio TOPBLOG 2010.
Sáb Jan 21, 2012 1:30 pm por Eduardo
Complexidade irredutível
conceito usado pelos proponentes do design inteligente segundo o qual certos sistemas biológicos possuem uma complexidade segundo a qual é altamente improvável que tenha surgido de forma evolutiva a partir de predecessores mais simples, ou "menos completos", através de mutações aleatórias vantajosas e seleção natural ocorridas naturalmente, i.e. sem a interferência de inteligência, pois tais sistemas biológicos só poderiam ser funcionais se todas as suas partes estivessem presentes e montadas na ordem certa. É um dos dois principais argumentos usados para apoiar o design inteligente, o outro sendo a complexidade especificada.[1] O conceito de complexidade irredutível é majoritariamente descartado pela comunidade científica[2] e bem assim a proposta do design inteligente na qual é constantemente utilizado para suportar, que é considerado globalmente como pseudociência.[3]
O bioquímico, professor Michael Behe, autor do argumento da complexidade irredutível, define um sistema irredutivelmente complexo como um sistema "composto de várias partes bem combinadas que interagem e que contribuem para a função básica do sistema, onde a remoção de qualquer das partes faz com que o sistema pare de funcionar".[4] Ele aponta alguns exemplos que, segundo ele, demonstram que as formas biológicas modernas não poderiam ter evoluído naturalmente. Os críticos consideram que a maioria, ou todos os exemplos são baseados em uma compreensão errônea do funcionamento dos sistemas biológicos em questão, e consideram a baixa qualidade dos exemplos uma prova excelente do argumento pela ignorância. No processo de 2005, Kitzmiller v. Dover Area School District, Behe testemunhou sobre a complexidade irredutível. A corte entendeu que "a alegação de complexidade irredutível do professor Behe foi refutada em trabalhos de pesquisa revisados por pares, e foi rejeitado pela comunidade científica em geral".[5] Apesar disso, a complexidade irredutível continua sendo citada como um argumento importante pelos criacionistas, particularmente pelos proponentes do design inteligente.
Recentemente, biólogos conseguiram determinar parte do processo evolutivo dos olhos, acabando com a teoria da complexidade irredutível, que usa estas estruturas como seu exemplo básico.[6]
Definições
O termo "complexidade irredutível" foi definido originalmente por Behe como sendo:
Uma segunda definição dada por Behe (sua "definição evolucionária") é esta:
Predecessores
O argumento da complexidade irredutível é um descendente do argumento teleológico para Deus (o argumento do projeto ou da complexidade). Ele diz que por que certas coisas na natureza são muito complicadas, elas devem ter sido projetadas. William Paley tornou-se famoso por declarar, em sua analogia do relojoeiro, de 1802, que a complexidade na natureza implica um Deus pela mesma razão que a existência de um relógio implica a existência de um relojoeiro.[7] Este argumento é bem antigo, e pode ser encontrado até na obra de Cícero, De natura deorum, ii.34.[8][9]
Até o século XVIII
Galeno (séculos I e II d. C.) escreveu sobre o grande número de partes no corpo e seus relacionamentos, cuja observação foi citada como evidência da criação.[10] A ideia que especificamente a interdependências entre partes pode ter implicações para a origem dos seres vivos foi levantada por autores, começando com Pierre Gassendi no meio do século XVII[11] e John Wilkins, que escreveu (citando Galeno), "Agora imaginar que todas estas coisas, de acordo com seus vários tipos, podem ser colocadas em sua forma e ordem regular, para a qual um número infinito de Intenções são necessárias, sem o projeto de um Agente inteligente, deve ser irracional no mais alto grau."[12] Ao fim do século XVII Thomas Burnet referiu-se a "uma multitude de peças apropriadamente reunidas" para argumentar contra a eternidade da vida.[13] No início do século XVII, Nicolas Malebranche[14] escreveu "Um corpo organizado contém uma infinidade de partes que dependem mutuamente umas das outras em relação a finalidades particulares, todas as quais devem estar formadas para funcionar como um todo," argumentando a favor da preformação, em vez da epigênese, do indivíduo. Um argumento semelhante sobre as origens do indivíduo foi feita por outros estudantes de história natural do século XVIII.
Século XIX
Quando fazemos a transição para o século XIX, começam a aparecer as referências à evolução.
O capítulo XV da Natural Theology (Teologia Natural) de Paley discute extensivamente o que ele chamou de "relações" das partes dos seres vivos como uma indicação de seu design.[7] Em uma aplicação diferente, no início do século XIX, Georges Cuvier usou o conceito de "correlação das partes" ao estabelecer a anatomia de animais a partir de restos fragmentados.[15][16]
Apesar de não ter dado origem ao termo, Charles Darwin identificou o argumento como uma forma possível de falsear a predição da teoria da evolução desde o início. Em "A Origem das Espécies", ele escreveu "se for possível demonstrar que qualquer órgão complexo exista, que não possa ter sido formado por pequenas mutações numerosas e sucessivas, minha teoria estará destruída. Mas eu não consigo encontrar tal exemplo."[17] A teoria da evolução de Darwin desafia o argumento teleológico ao postular uma explicação alternativa à do projetista inteligente, a evolução pela seleção natural. O argumento a partir da complexidade irredutível tenta demonstrar que certas funções biológicas não podem ser o resultado da evolução Darwiniana.
Século XX
Hermann Muller, no início do século XX, discutiu um conceito similar à complexidade irredutível. Entretanto, longe de a ver como um problema para a evolução, Muller descreveu a "interdependência" de funções biológicas como uma consequência esperada da evolução, que levaria à irreversibilidade de algumas mudanças evolucionárias.[18] Ele escreveu "Estando finalmente entrelaçado, como está, no tecido mais íntimo do organismo, o novo caráter não pode ser descartado com impunidade, e se torna vitalmente necessário."[19]
Em 1974, o Criacionista da Terra Jovem Henry M. Morris introduziu um conceito similar em seu livro "Criacionismo Científico" no qual ele escreve: "este especto pode ser atacado quantitativamente, usando princípios simples da probabilidade matemática. O problema é simplesmente se pode um sistema complexo, no qual muitos componentes funcionam unidos, e no qual cada componente é individualmente necessário para o funcionamento eficiente do todo, ter surgido por processos aleatórios."[20]
Em 1981, Ariel Roth, na defesa da posição da ciência da criação no caso de McLean vs. Arkansas, disse das "estruturas complexas integradas" que "Este sistema não pode ser funcional a menos que todas as partes estejam ali… Como estas partes sobreviveram durante a evolução…?"[21]
Em 1985 Cairns-Smith escreveu sobre a "interconexão", "Como pode uma colaboração complexa entre componentes evoluir em pequenos passos?" e usou a analogia da estrutura chamada centralização, usada para construir um arco, sendo removida depois: "Com certeza houve 'centralização'. Antes que os múltiplos componentes da bioquímica atual possa curvar-se sozinha ela deve primeiro curvar-se sobre outra coisa."[22] Entretanto, nem Muller nem Cairns-Smith alegaram que suas idéias fossem evidência de algo sobrenatural.[23]
Uma dissertação apoiando o criacionismo publicada em 1994 referiu-se aos flagelos bacterianos como apresentando "componentes múltiplos e integrados", onde "nada neles funciona a menos que todos seus complexos componentes estejam juntos" e solicitou ao leitor que "imagine os efeitos da seleção natural nestes organismos que tenham fortuitamente evoluído o flagelo… sem os mecanismos de controle concomitantes [sic]".[24][25]
Um conceito anterior de sistemas irredutivelmente complexos vem de Ludwig von Bertalanffy, um biólogo austríaco do século XX.[26] Ele acreditava que sistemas complexos deveriam ser examinados como sistemas completos e irredutíveis para se compreender como funcionam. Ele estendeu seu trabalho sobre complexidade biológica em uma teoria geral de sistemas em um livro chamado General System Theory (Teoria Geral de Sistemas). Após James Watson e Francis Crick terem publicado a estrutura do DNA no início da década de 1950, a Teoria Geral de Sistemas perdeu muitos de seus seguidores nas ciências físicas e biológicas.[27] No entanto, a teoria dos Sistemas permaneceu popular nas ciências sociais muito tempo depois de ter sido abandonada nas ciências físicas e biológicas.
Origens
Michael Behe desenvolveu suas idéias sobre este conceito em torno de 1992, nos primeiros dias do 'movimento da cunha', e apresentou suas idéias pela primeira vez sobre a "complexidade irredutível" em junho de 1993 quando o "grupo de professores Johnson-Behe" se reuniu em Pajaro Dunes na Califórnia.[28] Ele delineou suas idéias na segunda edição do livro "Of Pandas and People" publicado em 1993, extensivamente revisando o capítulo 6, "Biochemical Similarities" (Similaridades Bioquímicas) com novas seções no mecanismo complexo da coagulação do sangue e a origem das proteínas.[29]
Ele usou o termo "complexidade irredutível" pela primeira vez em seu livro de 1996, "Darwin's Black Box", para referir-se a certos sistemas bioquímicos celulares complexos. Ele declara que mecanismos evolucionários não podem explicar o desenvolvimento de sistemas com tal "complexidade irredutível". notadamente, Behe dá o crédito ao filósofo William Paley pelo conceito original, e não von Bertalanffy, e sugere que sua aplicação do conceito a sistemas biológicos é completamente original. Defensores do design inteligente alegam que os sistemas de complexidade irredutível foram deliberadamente elaborados por alguma forma de inteligência.
Em 2001 Michael Behe escreveu: "Há uma assimetria entre minha definição atual de complexidade irredutível e a tarefa que a seleção natural deve fazer. Eu espero reparar este defeito em trabalhos futuros." Behe explicou especificamente que "a definição atual coloca o foco na remoção de uma parte de um sistema que já esteja funcionando", mas que a "tarefa difícil que a evolução Darwiniana encara, entretanto, não é remover partes de sistemas sofisticados pre-existentes; deve ser colocar juntos componentes que fazem um novo sistema em primeiro lugar."[30] No julgamento de 2005 de Kitzmiller v. Dover Area School District, Behe testemunhou sob juramento que ele "não julga [a assimetria] séria o suficiente para [ter revisado o livro] até então."[31]
Além disso, Behe testemunhou que a presença de complexidade irredutível em organismos não descartaria o envolvimento de mecanismos evolucionários no desenvolvimento da vida orgânica. Ele também testemunhou que não sabia de "artigos revisados por pares em publicações científicas discutindo o design inteligente da cascata da coagulação sanguínea," mas que havia "provavelmente um grande número de artigos revisados por pares em publicações científicas que demonstram que o sistema de coagulação do sangue é de fato um arranjo significativo de partes de grande complexidade e sofisticação."[32] (O resultado do julgamento foi a sentença que "o design inteligente não é ciência e é essencialmente religioso em sua natureza").[33]
De acordo com a teoria da evolução, as variações genéticas acontecem sem um projeto ou intenção específica. O ambiente "seleciona" as variações que são mais aptas, que são então passadas à próxima geração dos organismos. As mudanças acontecem pela operação gradual das forças naturais durante o tempo, algumas vezes lentamente, outras mais rápido (veja o equilíbrio pontuado). Este processo é capaz de adaptar estruturas complexas a partir de estruturas iniciais simples, ou converter estruturas complexas de uma função a outra (veja en:spandrel). A maioria dos defensores do design inteligente aceitam que a evolução aconteça através da mutação e seleção natural ao "nível micro", como a mudança da frequência relativa dos diversos comprimentos de bico dos tentilhões, mas alega que ela não pode explicar a complexidade irredutível, porque nenhuma das partes de um sistema irredutível seria funcional ou vantajosa até que o sistema completo estivesse formado.
[editar] A analogia da ratoeira


Michael Behe acredita que muitos aspectos da vida apresentam evidências de design, usando a ratoeira em uma analogia que é criticada por outros[34]
Behe usa a ratoeira como um exemplo ilustrativo deste conceito. Uma ratoeira consiste de várias partes que interagem - a base, o prendedor, a mola, o martelo. Behe alega que todas as partes devem estar presentes para que a ratoeira funcione, e que a remoção de qualquer destas peças destrói a função da ratoeira. De forma semelhante, sistemas biológicos precisam da ação em conjunto de múltiplas partes para funcionar. Os defensores do design inteligente alegam que a seleção natural não poderia criar do nada estes sistemas para os quais a ciência não encontrou um caminho evolucionário de pequenas modificações sucessivas, por que a função selecionada só está presente quando todas as partes estão montadas. Os exemplos originais de Behe de mecanismos de complexidade irredutível incluem o flagelo bacteriano da Escherichia coli, a cascata da coagulação do sangue, cílios, e o sistema imunitário adaptativo.
Behe alega que os órgãos e funções biológica que são irredutivelmente complexos não podem ser completamente explicados pelos modelos atuais da evolução. Ele alega que:
A complexidade irredutível não é uma alegação que a evolução não acontece, mas uma alegação que ela é "incompleta". No último capítulo do livro Darwin's Black Box, Behe explica sua visão de que a complexidade irredutível é a prova do design inteligente. Os principais críticos, entretanto, alegam que a complexidade irredutível, como definida por Behe, pode ser gerada por mecanismos evolucionários conhecidos. A alegação de Behe de que nenhuma literatura científica modelou apropriadamente as origens dos sistemas bioquímicos através de mecanismos evolucionários foi refutada pelo TalkOrigins. O juiz no caso de Dover escreveu "Ao definir a complexidade irredutível na forma que o fez, o Professor Behe tenta excluir o fenômeno da exaptação por uma definição, ignorando desta forma abundantes evidências que refutam seu argumento. Notavelmente, o NAS rejeitou a alegação de Behe de complexidade irredutível…"
[editar] Exemplos apresentados
Behe e outros sugeriram que um certo número de aspectos biológicos que eles acreditam ser irredutivelmente complexos.
[editar] Ciclo de Krebs: Início-Fim Interligados
O ciclo do ácido cítrico começa com o Acetil-CoA, transferindo seu grupo acetila de dois carbonos ao composto receptor oxaloacetato, de quatro carbonos, formando um composto de seis carbonos, o citrato.
O citrato então passa por uma série de transformações químicas, perdendo dois grupos carboxila na forma de CO2. Os carbonos liberados na forma de CO2 são oriundos do oxaloacetato, e não diretamente do Acetil-CoA. Os carbonos doados pelo Acetil-CoA se tornam parte do oxaloacetato após o primeiro passo do ciclo do ácido cítrico.
A transformação dos carbonos doados pelo Acetil-CoA em CO2 requer vários passos no ciclo de Krebs. No entanto, por causa do papel do ácido cítrico no anabolismo (síntese de substâncias orgânicas), ele pode não ser perdido já que muitas substâncias intermediárias do ciclo também são usadas como precursoras para a biosíntese em outras moléculas.
A maior parte da energia disponível graças ao processo oxidativo do ciclo é transferida por elétrons altamente energéticos que reduzem o NAD+, tranformando-o em NADH. Para cada grupo acetila que entra no ciclo de Krebs, três moléculas de NADH são produzidas (o equivalente a 2,5 ATPs).
Elétrons também são transferidos ao receptor Q, formando QH2.
No final de cada ciclo, o Oxoalocetato de quatro carbonos é regenerado, e o processo continua utilizando o resultado do último processo como base principal para se obter um novo ciclo [35]
[editar] Cascata de coagulação sanguínea
A coagulação sanguínea ou a cascata de coagulação nos vertebrados é um caminho biológico complexo que é apresentado como um exemplo de aparente complexidade irredutível.
O argumento da complexidade irredutível assume que as partes necessárias de um sistema sempre são necessárias, e portando não podem ter sido acrescido sequencialmente. Entretanto, na evolução, algo que é meramente vantajoso a princípio pode mais tarde se tornar necessário. Muitas estruturas chamadas irredutíveis foram encontradas em outros organismos como sistemas muito mais simples que utilizam menos partes. Estes sistemas, por sua vez, podem ter tido precursores até mais simples que agora estão extintos. O "argumento da improbabilidade" também apresenta de forma errada a seleção natural. É correto dizer que um conjunto de mutações simultâneas que formam uma proteína de estrutura complexa é improvável, mas não foi isto que Darwin defendia. Sua explicação é baseada em pequenas mudanças acumuladas que acontecem sem um objetivo final. Cada passo tem que ser vantajoso por si mesmo, apesar de os biólogos não consigam entender a razão por trás de todos eles.
[editar] Olho
O olho é um exemplo famoso de uma estrutura irredutivelmente complexa, devido a suas muitas partes elaboradas e interconectadas, parecendo todas elas dependentes uma da outra. O olho é frequentemente citado por defensores do design inteligente e criacionismo como um exemplo de complexidade irredutível. Behe usou o "problema do desenvolvimento do olho" como uma evidência do design inteligente no livro Darwin's Black Box. Apesar de Behe reconhecer que a evolução de grandes estruturas anatômicas do olho serem bem explicadas, ele alegou que a complexidade das reações bioquímicas necessárias a nível molecular para a sensibilidade à luz ainda desafiam as explicações. O criacionista Jonathan Sarfati descreveu o olho como o "grande desafio" dos biólogos evolucionistas e "um exemplo de soberba 'complexidade irredutível' na criação de Deus", apontando especificamente para sua suposta "vasta complexidade" necessária para a transparência.
Em uma passagem geralmente citada de forma errada de "A Origem das Espécies", Charles Darwin parece reconhecer o desenvolvimento do olho como uma dificuldade para sua teoria. Entretanto, a citação no devido contexto mostra que Darwin possuía uma boa compreensão da evolução do olho. Ele aponta que "supor que o olho [...] possa ter sido formado pela seleção natural parece, eu confesso frequentemente, absurdo no mais alto grau". Entretanto, esta observação era apenas um recurso retórico de Darwin. Ele prossegue explicando que se a evolução gradual do olho for demonstrada como possível, "a dificuldade de acreditar que um olho perfeito e complexo possa ser formado pela seleção natural [...] dificilmente pode ser considerada real". Ele então seguiu apresentando um provável roteiro de evolução do olho usando exemplos de olhos gradualmente mais complexos, de várias espécies.
Desde os dias de Darwin, a ancestralidade do olho é bem melhor compreendida. Apesar de que o entendimento da construção dos olhos antigos através do registro fóssil ser problemático devido ao fato dos tecidos macios não deixarem impressões ou restos, a evidência genética e de anatomia comparativa tem apoiado a idéia de uma ancestralidade comum a todos os olhos.
Conforme Behe admite, a evidência atual sugere possíveis linhas evolucionárias para as origens das estruturas anatômicas do olho, por exemplo, que os olhos tiveram origem como simples manchas de células fotorreceptoras que podem detectar a presença ou ausência de luz, mas não sua direção. Ao desenvolver uma pequena depressão para as células fotossensitivas, os organismos ganharam uma melhor capacidade de descobrir a origem da luz, e como o contínuo aprofundamento da depressão em um buraco de forma que a luz atinja certas células dependendo do seu ângulo, uma informação visível cada vez mais precisa se tornou possível. A abertura do olho era então diminuída e forma que a luz se tornou focada, transformando o olho em uma câmera pinhole e permitindo ao organismo perceber formas — o náutilo é um exemplo moderno de um animal com um olho deste tipo. Finalmente, a camada protetiva de células transparentes sobre a abertura se diferenciou em uma lente primitiva, e o interior do olho foi preenchido com humores para ajudar a colocar as imagens em foco. Desta forma, os biólogos modernos reconhecem os olhos como uma estrutura relativamente simples que evoluiu, e muitos dos principais desenvolvimentos da evolução do olho crê-se terem acontecido em alguns poucos milhões de anos, durante a explosão cambriana. Entretanto, de acordo com Behe, a complexidade da sensibilidade à luz no nível molecular e as reações bioquímicas necessárias para as "primeiras manchas simples de fotoreceptores" ainda desafiam as explicações.
[editar] Flagelo
O flagelo de certas bactérias constitui um motor molecular exigindo a interação de cerca de 40 partes proteicas complexas, e a ausência de qualquer uma destas proteínas faz com que o flagelo não funcione. Behe mantém que o "mecanismo" do flagelo é irredutivelmente complexo por que se tentarmos reduzir sua complexidade colocando um estágio anterior mais simples de seu desenvolvimento evolucionário, obteremos um organismo que não funciona de maneira apropriada.
Os cientistas entendem que este argumento já foi refutado à luz de pesquisas recentes. Eles apontam que o corpo basal do flagelo é similar ao sistema secretor Tipo III (TTSS na sigla em inglês), uma estrutura em forma de agulha que alguns germes patogênicos como a Salmonella e a Yersinia pestis usam para injetar toxinas em células eucariontes vivas. A base da agulha possui dez elementos em comum com o flagelo, mas ainda faltam quarenta das proteínas que fazem o flagelo funcional. Entretanto, este sistema parece negar a alegação de que tirar qualquer das partes do flagelo o transformam em uma estrutura inútil. Isto fez com que Kenneth Miller notasse que "As partes destes sistema supostamente irredutivelmente complexo na verdade tenham funções próprias."
Referências
↑ Muller HJ. (1918). "Genetic variability, twin hybrids and constant hybrids, in a case of balanced lethal factors". Genetics 3 (5): 422–99., especially pages 463–4.
↑ Muller HJ. (1939). "Reversibility in evolution considered from the standpoint of genetics". Biological Reviews of the Cambridge Philosophical Society 4 (3): 261–80, quotation from 272.
↑ Morris, Henry. Scientific creationism. 2nd ed. San Diego, Calif: Creation-Life Publishers, 1974. 59 p. ISBN 0-89051-003-2
↑ Keough, Mark J.; Geisler, Norman L.. The Creator in the courtroom "Scopes II": the 1981 Arkansas creation-evolution trial. Milford, Mich: Mott Media, 1982. p. 146. ISBN 0-88062-020-X
↑ Cairns-Smith, A. G.. Seven clues to the origin of life: a scientific detective story. Cambridge, UK: Cambridge University Press, 1986. 39, 58–64 p. ISBN 0-521-27522-9
↑ Mark Perakh (2008). Bacteria Flagella Look Like Man-made Machines. Skeptic (U.S. magazine). Página visitada em 2008-12-06.
↑ Lumsden RD. (June 1994). "Not So Blind A Watchmaker". Creation Research Society Quarterly 31 (1): 13–22, quotations from 13, 20.
↑ Scott EC, Matzke NJ. (May 2007). "Biological design in science classrooms". Proc Natl Acad Sci USA. 104 Suppl 1: 8669–76, See page 8672. DOI:10.1073/pnas.0701505104. PMID 17494747.
↑ Ludwig von Bertalanffy (1952). Problems of Life: An Evaluation of Modern Biological and Scientific Thought, pg 148 ISBN 1-131-79242-4
↑ Monod, Jacques. Chance and necessity: an essay on the natural philosophy of modern biology. New York: Vintage Books, 1972. ISBN 0-394-71825-9
↑ Barbara Forrest, The Wedge at Work. Talk Reason.
Forrest B. In: Pennock RT. Intelligent design creationism and its critics: philosophical, theological, and scientific perspectives. Cambridge, Mass: MIT Press, 2001. 5–54 p. ISBN 0-262-66124-1
↑ The New Pandas: Has Creationist Scholarship Improved? Comments on 1993 Revisions by Frank J. Sonleitner (1994)
Introduction: Of Pandas and People, the foundational work of the 'Intelligent Design' movement by Nick Matzke 2004,
Design on Trial in Dover, Pennsylvania by Nicholas J Matzke, NCSE Public Information Project Specialist
↑ Behe MJ. (November 2001). "Reply to My Critics: A Response to Reviews of Darwin's Black Box: The Biochemical Challenge to Evolution". Biology and Philosophy 16 (5): 685–709. DOI:10.1023/A:1012268700496.
↑ Behe's testimony in Kitzmiller v. Dover
↑ Behe, Michael 2005 Kitzmiller v. Dover Area School District 4: whether ID is science (p. 88)
↑ Kitzmiller v. Dover Area School District 6: Conclusion, section H
↑ A reducibly complex mousetrap (graphics-intensive, requires JavaScript)
↑ UFP: A lógica do Ciclo de Krebs
conceito usado pelos proponentes do design inteligente segundo o qual certos sistemas biológicos possuem uma complexidade segundo a qual é altamente improvável que tenha surgido de forma evolutiva a partir de predecessores mais simples, ou "menos completos", através de mutações aleatórias vantajosas e seleção natural ocorridas naturalmente, i.e. sem a interferência de inteligência, pois tais sistemas biológicos só poderiam ser funcionais se todas as suas partes estivessem presentes e montadas na ordem certa. É um dos dois principais argumentos usados para apoiar o design inteligente, o outro sendo a complexidade especificada.[1] O conceito de complexidade irredutível é majoritariamente descartado pela comunidade científica[2] e bem assim a proposta do design inteligente na qual é constantemente utilizado para suportar, que é considerado globalmente como pseudociência.[3]

O bioquímico, professor Michael Behe, autor do argumento da complexidade irredutível, define um sistema irredutivelmente complexo como um sistema "composto de várias partes bem combinadas que interagem e que contribuem para a função básica do sistema, onde a remoção de qualquer das partes faz com que o sistema pare de funcionar".[4] Ele aponta alguns exemplos que, segundo ele, demonstram que as formas biológicas modernas não poderiam ter evoluído naturalmente. Os críticos consideram que a maioria, ou todos os exemplos são baseados em uma compreensão errônea do funcionamento dos sistemas biológicos em questão, e consideram a baixa qualidade dos exemplos uma prova excelente do argumento pela ignorância. No processo de 2005, Kitzmiller v. Dover Area School District, Behe testemunhou sobre a complexidade irredutível. A corte entendeu que "a alegação de complexidade irredutível do professor Behe foi refutada em trabalhos de pesquisa revisados por pares, e foi rejeitado pela comunidade científica em geral".[5] Apesar disso, a complexidade irredutível continua sendo citada como um argumento importante pelos criacionistas, particularmente pelos proponentes do design inteligente.
Recentemente, biólogos conseguiram determinar parte do processo evolutivo dos olhos, acabando com a teoria da complexidade irredutível, que usa estas estruturas como seu exemplo básico.[6]
O termo "complexidade irredutível" foi definido originalmente por Behe como sendo:
Defensores do design inteligente usam esta expressão para se referir a sistemas biológicos e órgãos que eles acreditam não poderem ter surgido por qualquer série de pequenas mudanças. Eles alegam que qualquer coisa menos que a forma completa do sistema o órgão completo não funcionaria, ou seria de fato danoso ao organismo, que portanto não poderia sobreviver ao processo de seleção natural. Apesar deles aceitarem que alguns órgãos e sistemas complexos podem ser explicados pela evolução, alegam que algumas funções biológicas e órgãos que são irredutivelmente complexos não podem ser explicados pelos modelos atuais, e que um projetista inteligente deve ter criado a vida ou guiado sua evolução. Portanto, o debate sobre a complexidade irredutível se reduz a duas questões: se a complexidade irredutível pode ser encontrada na natureza, e qual a significância se ela existir na natureza.
Um sistema único que é composto de várias partes que interagem e contribuem para a função básica, e onde a remoção de qualquer uma das partes faz com que o sistema efetivamente pare de funcionar.– (Darwin's Black Box p39 in the 2006 edition)
Uma segunda definição dada por Behe (sua "definição evolucionária") é esta:
William Dembski, outro defensor do design inteligente, dá esta definição:
Um caminho de complexidade irredutível é um que contém um ou mais passos não selecionados (ou seja, uma ou mais mutações necessárias-mas-não-selecionadas). O grau de complexidade irredutível é o número de passos não selecionados no caminho.
Histórico
Um sistema que executa uma função básica é irredutivelmente complexo se ele inclui um conjunto de partes individualizadas de forma não arbitrária, bem-combinadas, que interagem mutuamente, de forma que cada parte no conjunto é indispensável para manter a função básica, e portanto original, do sistema. O conjunto destas partes indispensáveis é conhecido como o núcleo irredutível do sistema.– No Free Lunch: Why Specified Complexity Cannot Be Purchased without Intelligence. by William Dembski pp. 285
Predecessores
O argumento da complexidade irredutível é um descendente do argumento teleológico para Deus (o argumento do projeto ou da complexidade). Ele diz que por que certas coisas na natureza são muito complicadas, elas devem ter sido projetadas. William Paley tornou-se famoso por declarar, em sua analogia do relojoeiro, de 1802, que a complexidade na natureza implica um Deus pela mesma razão que a existência de um relógio implica a existência de um relojoeiro.[7] Este argumento é bem antigo, e pode ser encontrado até na obra de Cícero, De natura deorum, ii.34.[8][9]
Até o século XVIII
Galeno (séculos I e II d. C.) escreveu sobre o grande número de partes no corpo e seus relacionamentos, cuja observação foi citada como evidência da criação.[10] A ideia que especificamente a interdependências entre partes pode ter implicações para a origem dos seres vivos foi levantada por autores, começando com Pierre Gassendi no meio do século XVII[11] e John Wilkins, que escreveu (citando Galeno), "Agora imaginar que todas estas coisas, de acordo com seus vários tipos, podem ser colocadas em sua forma e ordem regular, para a qual um número infinito de Intenções são necessárias, sem o projeto de um Agente inteligente, deve ser irracional no mais alto grau."[12] Ao fim do século XVII Thomas Burnet referiu-se a "uma multitude de peças apropriadamente reunidas" para argumentar contra a eternidade da vida.[13] No início do século XVII, Nicolas Malebranche[14] escreveu "Um corpo organizado contém uma infinidade de partes que dependem mutuamente umas das outras em relação a finalidades particulares, todas as quais devem estar formadas para funcionar como um todo," argumentando a favor da preformação, em vez da epigênese, do indivíduo. Um argumento semelhante sobre as origens do indivíduo foi feita por outros estudantes de história natural do século XVIII.
Século XIX
Quando fazemos a transição para o século XIX, começam a aparecer as referências à evolução.
O capítulo XV da Natural Theology (Teologia Natural) de Paley discute extensivamente o que ele chamou de "relações" das partes dos seres vivos como uma indicação de seu design.[7] Em uma aplicação diferente, no início do século XIX, Georges Cuvier usou o conceito de "correlação das partes" ao estabelecer a anatomia de animais a partir de restos fragmentados.[15][16]
Apesar de não ter dado origem ao termo, Charles Darwin identificou o argumento como uma forma possível de falsear a predição da teoria da evolução desde o início. Em "A Origem das Espécies", ele escreveu "se for possível demonstrar que qualquer órgão complexo exista, que não possa ter sido formado por pequenas mutações numerosas e sucessivas, minha teoria estará destruída. Mas eu não consigo encontrar tal exemplo."[17] A teoria da evolução de Darwin desafia o argumento teleológico ao postular uma explicação alternativa à do projetista inteligente, a evolução pela seleção natural. O argumento a partir da complexidade irredutível tenta demonstrar que certas funções biológicas não podem ser o resultado da evolução Darwiniana.
Século XX
Hermann Muller, no início do século XX, discutiu um conceito similar à complexidade irredutível. Entretanto, longe de a ver como um problema para a evolução, Muller descreveu a "interdependência" de funções biológicas como uma consequência esperada da evolução, que levaria à irreversibilidade de algumas mudanças evolucionárias.[18] Ele escreveu "Estando finalmente entrelaçado, como está, no tecido mais íntimo do organismo, o novo caráter não pode ser descartado com impunidade, e se torna vitalmente necessário."[19]
Em 1974, o Criacionista da Terra Jovem Henry M. Morris introduziu um conceito similar em seu livro "Criacionismo Científico" no qual ele escreve: "este especto pode ser atacado quantitativamente, usando princípios simples da probabilidade matemática. O problema é simplesmente se pode um sistema complexo, no qual muitos componentes funcionam unidos, e no qual cada componente é individualmente necessário para o funcionamento eficiente do todo, ter surgido por processos aleatórios."[20]
Em 1981, Ariel Roth, na defesa da posição da ciência da criação no caso de McLean vs. Arkansas, disse das "estruturas complexas integradas" que "Este sistema não pode ser funcional a menos que todas as partes estejam ali… Como estas partes sobreviveram durante a evolução…?"[21]
Em 1985 Cairns-Smith escreveu sobre a "interconexão", "Como pode uma colaboração complexa entre componentes evoluir em pequenos passos?" e usou a analogia da estrutura chamada centralização, usada para construir um arco, sendo removida depois: "Com certeza houve 'centralização'. Antes que os múltiplos componentes da bioquímica atual possa curvar-se sozinha ela deve primeiro curvar-se sobre outra coisa."[22] Entretanto, nem Muller nem Cairns-Smith alegaram que suas idéias fossem evidência de algo sobrenatural.[23]
Uma dissertação apoiando o criacionismo publicada em 1994 referiu-se aos flagelos bacterianos como apresentando "componentes múltiplos e integrados", onde "nada neles funciona a menos que todos seus complexos componentes estejam juntos" e solicitou ao leitor que "imagine os efeitos da seleção natural nestes organismos que tenham fortuitamente evoluído o flagelo… sem os mecanismos de controle concomitantes [sic]".[24][25]
Um conceito anterior de sistemas irredutivelmente complexos vem de Ludwig von Bertalanffy, um biólogo austríaco do século XX.[26] Ele acreditava que sistemas complexos deveriam ser examinados como sistemas completos e irredutíveis para se compreender como funcionam. Ele estendeu seu trabalho sobre complexidade biológica em uma teoria geral de sistemas em um livro chamado General System Theory (Teoria Geral de Sistemas). Após James Watson e Francis Crick terem publicado a estrutura do DNA no início da década de 1950, a Teoria Geral de Sistemas perdeu muitos de seus seguidores nas ciências físicas e biológicas.[27] No entanto, a teoria dos Sistemas permaneceu popular nas ciências sociais muito tempo depois de ter sido abandonada nas ciências físicas e biológicas.
Origens
Michael Behe desenvolveu suas idéias sobre este conceito em torno de 1992, nos primeiros dias do 'movimento da cunha', e apresentou suas idéias pela primeira vez sobre a "complexidade irredutível" em junho de 1993 quando o "grupo de professores Johnson-Behe" se reuniu em Pajaro Dunes na Califórnia.[28] Ele delineou suas idéias na segunda edição do livro "Of Pandas and People" publicado em 1993, extensivamente revisando o capítulo 6, "Biochemical Similarities" (Similaridades Bioquímicas) com novas seções no mecanismo complexo da coagulação do sangue e a origem das proteínas.[29]
Ele usou o termo "complexidade irredutível" pela primeira vez em seu livro de 1996, "Darwin's Black Box", para referir-se a certos sistemas bioquímicos celulares complexos. Ele declara que mecanismos evolucionários não podem explicar o desenvolvimento de sistemas com tal "complexidade irredutível". notadamente, Behe dá o crédito ao filósofo William Paley pelo conceito original, e não von Bertalanffy, e sugere que sua aplicação do conceito a sistemas biológicos é completamente original. Defensores do design inteligente alegam que os sistemas de complexidade irredutível foram deliberadamente elaborados por alguma forma de inteligência.
Em 2001 Michael Behe escreveu: "Há uma assimetria entre minha definição atual de complexidade irredutível e a tarefa que a seleção natural deve fazer. Eu espero reparar este defeito em trabalhos futuros." Behe explicou especificamente que "a definição atual coloca o foco na remoção de uma parte de um sistema que já esteja funcionando", mas que a "tarefa difícil que a evolução Darwiniana encara, entretanto, não é remover partes de sistemas sofisticados pre-existentes; deve ser colocar juntos componentes que fazem um novo sistema em primeiro lugar."[30] No julgamento de 2005 de Kitzmiller v. Dover Area School District, Behe testemunhou sob juramento que ele "não julga [a assimetria] séria o suficiente para [ter revisado o livro] até então."[31]
Além disso, Behe testemunhou que a presença de complexidade irredutível em organismos não descartaria o envolvimento de mecanismos evolucionários no desenvolvimento da vida orgânica. Ele também testemunhou que não sabia de "artigos revisados por pares em publicações científicas discutindo o design inteligente da cascata da coagulação sanguínea," mas que havia "provavelmente um grande número de artigos revisados por pares em publicações científicas que demonstram que o sistema de coagulação do sangue é de fato um arranjo significativo de partes de grande complexidade e sofisticação."[32] (O resultado do julgamento foi a sentença que "o design inteligente não é ciência e é essencialmente religioso em sua natureza").[33]
De acordo com a teoria da evolução, as variações genéticas acontecem sem um projeto ou intenção específica. O ambiente "seleciona" as variações que são mais aptas, que são então passadas à próxima geração dos organismos. As mudanças acontecem pela operação gradual das forças naturais durante o tempo, algumas vezes lentamente, outras mais rápido (veja o equilíbrio pontuado). Este processo é capaz de adaptar estruturas complexas a partir de estruturas iniciais simples, ou converter estruturas complexas de uma função a outra (veja en:spandrel). A maioria dos defensores do design inteligente aceitam que a evolução aconteça através da mutação e seleção natural ao "nível micro", como a mudança da frequência relativa dos diversos comprimentos de bico dos tentilhões, mas alega que ela não pode explicar a complexidade irredutível, porque nenhuma das partes de um sistema irredutível seria funcional ou vantajosa até que o sistema completo estivesse formado.
[editar] A analogia da ratoeira

Michael Behe acredita que muitos aspectos da vida apresentam evidências de design, usando a ratoeira em uma analogia que é criticada por outros[34]
Behe usa a ratoeira como um exemplo ilustrativo deste conceito. Uma ratoeira consiste de várias partes que interagem - a base, o prendedor, a mola, o martelo. Behe alega que todas as partes devem estar presentes para que a ratoeira funcione, e que a remoção de qualquer destas peças destrói a função da ratoeira. De forma semelhante, sistemas biológicos precisam da ação em conjunto de múltiplas partes para funcionar. Os defensores do design inteligente alegam que a seleção natural não poderia criar do nada estes sistemas para os quais a ciência não encontrou um caminho evolucionário de pequenas modificações sucessivas, por que a função selecionada só está presente quando todas as partes estão montadas. Os exemplos originais de Behe de mecanismos de complexidade irredutível incluem o flagelo bacteriano da Escherichia coli, a cascata da coagulação do sangue, cílios, e o sistema imunitário adaptativo.
Behe alega que os órgãos e funções biológica que são irredutivelmente complexos não podem ser completamente explicados pelos modelos atuais da evolução. Ele alega que:
Um sistema complexo irredutível não pode ser produzido diretamente (ou seja, pelo aperfeiçoamento contínuo da função inicial, que continua a funcionar pelos mesmos mecanismos) por pequenas modificações sucessivas, por que qualquer precursor de um sistema irredutivelmente complexo que está sem uma parte é por definição não-funcional
A complexidade irredutível não é uma alegação que a evolução não acontece, mas uma alegação que ela é "incompleta". No último capítulo do livro Darwin's Black Box, Behe explica sua visão de que a complexidade irredutível é a prova do design inteligente. Os principais críticos, entretanto, alegam que a complexidade irredutível, como definida por Behe, pode ser gerada por mecanismos evolucionários conhecidos. A alegação de Behe de que nenhuma literatura científica modelou apropriadamente as origens dos sistemas bioquímicos através de mecanismos evolucionários foi refutada pelo TalkOrigins. O juiz no caso de Dover escreveu "Ao definir a complexidade irredutível na forma que o fez, o Professor Behe tenta excluir o fenômeno da exaptação por uma definição, ignorando desta forma abundantes evidências que refutam seu argumento. Notavelmente, o NAS rejeitou a alegação de Behe de complexidade irredutível…"
[editar] Exemplos apresentados
Behe e outros sugeriram que um certo número de aspectos biológicos que eles acreditam ser irredutivelmente complexos.
[editar] Ciclo de Krebs: Início-Fim Interligados
O ciclo do ácido cítrico começa com o Acetil-CoA, transferindo seu grupo acetila de dois carbonos ao composto receptor oxaloacetato, de quatro carbonos, formando um composto de seis carbonos, o citrato.
O citrato então passa por uma série de transformações químicas, perdendo dois grupos carboxila na forma de CO2. Os carbonos liberados na forma de CO2 são oriundos do oxaloacetato, e não diretamente do Acetil-CoA. Os carbonos doados pelo Acetil-CoA se tornam parte do oxaloacetato após o primeiro passo do ciclo do ácido cítrico.
A transformação dos carbonos doados pelo Acetil-CoA em CO2 requer vários passos no ciclo de Krebs. No entanto, por causa do papel do ácido cítrico no anabolismo (síntese de substâncias orgânicas), ele pode não ser perdido já que muitas substâncias intermediárias do ciclo também são usadas como precursoras para a biosíntese em outras moléculas.
A maior parte da energia disponível graças ao processo oxidativo do ciclo é transferida por elétrons altamente energéticos que reduzem o NAD+, tranformando-o em NADH. Para cada grupo acetila que entra no ciclo de Krebs, três moléculas de NADH são produzidas (o equivalente a 2,5 ATPs).
Elétrons também são transferidos ao receptor Q, formando QH2.
No final de cada ciclo, o Oxoalocetato de quatro carbonos é regenerado, e o processo continua utilizando o resultado do último processo como base principal para se obter um novo ciclo [35]
[editar] Cascata de coagulação sanguínea
A coagulação sanguínea ou a cascata de coagulação nos vertebrados é um caminho biológico complexo que é apresentado como um exemplo de aparente complexidade irredutível.
O argumento da complexidade irredutível assume que as partes necessárias de um sistema sempre são necessárias, e portando não podem ter sido acrescido sequencialmente. Entretanto, na evolução, algo que é meramente vantajoso a princípio pode mais tarde se tornar necessário. Muitas estruturas chamadas irredutíveis foram encontradas em outros organismos como sistemas muito mais simples que utilizam menos partes. Estes sistemas, por sua vez, podem ter tido precursores até mais simples que agora estão extintos. O "argumento da improbabilidade" também apresenta de forma errada a seleção natural. É correto dizer que um conjunto de mutações simultâneas que formam uma proteína de estrutura complexa é improvável, mas não foi isto que Darwin defendia. Sua explicação é baseada em pequenas mudanças acumuladas que acontecem sem um objetivo final. Cada passo tem que ser vantajoso por si mesmo, apesar de os biólogos não consigam entender a razão por trás de todos eles.
[editar] Olho
O olho é um exemplo famoso de uma estrutura irredutivelmente complexa, devido a suas muitas partes elaboradas e interconectadas, parecendo todas elas dependentes uma da outra. O olho é frequentemente citado por defensores do design inteligente e criacionismo como um exemplo de complexidade irredutível. Behe usou o "problema do desenvolvimento do olho" como uma evidência do design inteligente no livro Darwin's Black Box. Apesar de Behe reconhecer que a evolução de grandes estruturas anatômicas do olho serem bem explicadas, ele alegou que a complexidade das reações bioquímicas necessárias a nível molecular para a sensibilidade à luz ainda desafiam as explicações. O criacionista Jonathan Sarfati descreveu o olho como o "grande desafio" dos biólogos evolucionistas e "um exemplo de soberba 'complexidade irredutível' na criação de Deus", apontando especificamente para sua suposta "vasta complexidade" necessária para a transparência.
Em uma passagem geralmente citada de forma errada de "A Origem das Espécies", Charles Darwin parece reconhecer o desenvolvimento do olho como uma dificuldade para sua teoria. Entretanto, a citação no devido contexto mostra que Darwin possuía uma boa compreensão da evolução do olho. Ele aponta que "supor que o olho [...] possa ter sido formado pela seleção natural parece, eu confesso frequentemente, absurdo no mais alto grau". Entretanto, esta observação era apenas um recurso retórico de Darwin. Ele prossegue explicando que se a evolução gradual do olho for demonstrada como possível, "a dificuldade de acreditar que um olho perfeito e complexo possa ser formado pela seleção natural [...] dificilmente pode ser considerada real". Ele então seguiu apresentando um provável roteiro de evolução do olho usando exemplos de olhos gradualmente mais complexos, de várias espécies.
Desde os dias de Darwin, a ancestralidade do olho é bem melhor compreendida. Apesar de que o entendimento da construção dos olhos antigos através do registro fóssil ser problemático devido ao fato dos tecidos macios não deixarem impressões ou restos, a evidência genética e de anatomia comparativa tem apoiado a idéia de uma ancestralidade comum a todos os olhos.
Conforme Behe admite, a evidência atual sugere possíveis linhas evolucionárias para as origens das estruturas anatômicas do olho, por exemplo, que os olhos tiveram origem como simples manchas de células fotorreceptoras que podem detectar a presença ou ausência de luz, mas não sua direção. Ao desenvolver uma pequena depressão para as células fotossensitivas, os organismos ganharam uma melhor capacidade de descobrir a origem da luz, e como o contínuo aprofundamento da depressão em um buraco de forma que a luz atinja certas células dependendo do seu ângulo, uma informação visível cada vez mais precisa se tornou possível. A abertura do olho era então diminuída e forma que a luz se tornou focada, transformando o olho em uma câmera pinhole e permitindo ao organismo perceber formas — o náutilo é um exemplo moderno de um animal com um olho deste tipo. Finalmente, a camada protetiva de células transparentes sobre a abertura se diferenciou em uma lente primitiva, e o interior do olho foi preenchido com humores para ajudar a colocar as imagens em foco. Desta forma, os biólogos modernos reconhecem os olhos como uma estrutura relativamente simples que evoluiu, e muitos dos principais desenvolvimentos da evolução do olho crê-se terem acontecido em alguns poucos milhões de anos, durante a explosão cambriana. Entretanto, de acordo com Behe, a complexidade da sensibilidade à luz no nível molecular e as reações bioquímicas necessárias para as "primeiras manchas simples de fotoreceptores" ainda desafiam as explicações.
[editar] Flagelo
O flagelo de certas bactérias constitui um motor molecular exigindo a interação de cerca de 40 partes proteicas complexas, e a ausência de qualquer uma destas proteínas faz com que o flagelo não funcione. Behe mantém que o "mecanismo" do flagelo é irredutivelmente complexo por que se tentarmos reduzir sua complexidade colocando um estágio anterior mais simples de seu desenvolvimento evolucionário, obteremos um organismo que não funciona de maneira apropriada.
Os cientistas entendem que este argumento já foi refutado à luz de pesquisas recentes. Eles apontam que o corpo basal do flagelo é similar ao sistema secretor Tipo III (TTSS na sigla em inglês), uma estrutura em forma de agulha que alguns germes patogênicos como a Salmonella e a Yersinia pestis usam para injetar toxinas em células eucariontes vivas. A base da agulha possui dez elementos em comum com o flagelo, mas ainda faltam quarenta das proteínas que fazem o flagelo funcional. Entretanto, este sistema parece negar a alegação de que tirar qualquer das partes do flagelo o transformam em uma estrutura inútil. Isto fez com que Kenneth Miller notasse que "As partes destes sistema supostamente irredutivelmente complexo na verdade tenham funções próprias."
Referências
- ↑ Forrest, Barbara (May,2007) (PDF), Understanding the Intelligent Design Creationist Movement: Its True Nature and Goals. (Entendendo o Movimento Criacionista do Design Inteligente: Sua Natureza Verdadeira e Objetivos) A Position Paper from the Center for Inquiry, Office of Public Policy, Washington, D.C.: Center for Inquiry, Inc., http://www.centerforinquiry.net/uploads/attachments/intelligent-design.pdf, visitado em 2007-08-22 .
- ↑ "We therefore find that Professor Behe’s claim for irreducible complexity has been refuted in peer-reviewed research papers and has been rejected by the scientific community at large." Ruling, Judge John E. Jones III, Kitzmiller v. Dover Area School District
- ↑ "True in this latest creationist variant, advocates of so-called intelligent design… use more slick, pseudoscientific language. They talk about things like 'irreducible complexity'" —
Shulman, Seth. Undermining science: suppression and distortion in the Bush Administration. Berkeley: University of California Press, 2006. p. 13. ISBN 0-520-24702-7 "for most members of the mainstream scientific community, ID is not a scientific theory, but a creationist pseudoscience."
David Mu. (Fall 2005). "Trojan Horse or Legitimate Science: Deconstructing the Debate over Intelligent Design". Harvard Science Review 19 (1).
Perakh M. (2005 Summer). "Why Intelligent Design Isn't Intelligent — Review of: Unintelligent Design". Cell Biol Educ. 4 (2): 121–2. DOI:10.1187/cbe.05-02-0071.
Mark D. Decker. College of Biological Sciences, General Biology Program, University of Minnesota Frequently Asked Questions About the Texas Science Textbook Adoption Controversy "The Discovery Institute and ID proponents have a number of goals that they hope to achieve using disingenuous and mendacious methods of marketing, publicity, and political persuasion. They do not practice real science because that takes too long, but mainly because this method requires that one have actual evidence and logical reasons for one's conclusions, and the ID proponents just don't have those. If they had such resources, they would use them, and not the disreputable methods they actually use." - ↑ Darwin's Black Box: The Biochemical Challenge to Evolution, Michael Behe, 1996, quoted in Irreducible Complexity and Michael Behe (retrieved 8 January 2006)
- ↑ Ruling, Kitzmiller v. Dover Area School District, page 64
- ↑ A Fascinante evolução do olho. Scientific American Brasil (agosto de 2011). Página visitada em 2 de agosto de 2011. "O olho humano é tão complexo que sua origem provoca discussão entre criacionistas e defensores do desenho inteligente, que o têm como exemplo básico do que chamam de complexidade irredutível: um sistema que não funciona na ausência de quaisquer de seus componentes e, portanto, não poderia ter evoluído naturalmente de uma forma mais primitiva. [...] Ainda assim, recentemente biólogos fizeram avanços significativos no estudo da origem do olho [...]. Os resultados indicam que o tipo de olho comum entre os vertebrados se formou há menos de 100 milhões de anos [...]. [...] essas descobertas sepultam a tese da complexidade irredutível e apoiam a teoria da evolução. Explicam ainda porque o olho, longe de ser uma peça de maquinaria criada à perfeição, exibe falhas evidentes – “cicatrizes” da evolução."
- ↑ a b William Paley: Natural Theology; or, Evidences of the Existence and Attributes of the Deity. Collected from the Appearances of Nature 12th edition, 1809.
- ↑ On the Nature of the Gods, translated by Francis Brooks, London: Methuen, 1896..
- ↑ See Henry Hallam's Introduction to the Literature of Europe part iii chapter iii paragraph 26 footnote u
- ↑ De Formatione Foetus=The Construction of the Embryo, chapter 11 in Galen: Selected Works, translated by P. N. Singer, The World's Classics, Oxford, Oxford University Press, 1997 ISBN 978-019-282450-9. One 18th-century reference to Galen is David Hume Dialogues Concerning Natural Religion, 1779, Part 12, § 3, page 215. Also see Galen's De Usu Partium=On the Usefulness of the Parts of the Body, translated and edited by Margaret Tallmadge May, Ithaca, New York, Cornell University Press, 1968, especially book XVII.
- ↑ De Generatione Animalium, chapter III. Partial translation in: Howard B. Adelmann, Marcello Malpighi and the Evolution of Embryology Ithaca, New York, Cornell University Press, 1966, volume 2, pages 811-812.
- ↑ Of the Principles and Duties of Natural Religion, London, 1675, book I, chapter 6, page 82.
- ↑ The Sacred Theory of the Earth, 2nd edition, London: Walter Kettilby, 1691. Book I Chapter IV page 43.
- ↑ De la recherche de la verité 6.2.4, 6th edition, 1712. English translation The Search after Truth, translated and edited by Thomas M. Lennon and Paul J. Olscamp, Cambridge University Press, 1997, ISBN 0-521-58004-8. Second paragraph from the end of the chapter, on page 465.
- ↑ Andrew Pyle. In: Smith JH. The problem of animal generation in early modern philosophy. Cambridge, UK: Cambridge University Press, 2006. 202–3 p. ISBN 0-521-84077-5
- ↑ The Chicken or the Egg.
- ↑ Darwin, Charles (1859). On the Origin of Species. London: John Murray.
- page 189, Chapter VI
Forrest B. In: Pennock RT. Intelligent design creationism and its critics: philosophical, theological, and scientific perspectives. Cambridge, Mass: MIT Press, 2001. 5–54 p. ISBN 0-262-66124-1
Introduction: Of Pandas and People, the foundational work of the 'Intelligent Design' movement by Nick Matzke 2004,
Design on Trial in Dover, Pennsylvania by Nicholas J Matzke, NCSE Public Information Project Specialist
» Mapa metabólico e complexidade irredutível
» Nobel discorda da tese sobre mudanças climáticas
» Em busca de uma nova teoria que considere a complexidade biológica
» A complexidade tecnológica da celula da qual Darwin era totalmente ignorante
» E então a “tese” da gravidade criadora de Stephen Hawking era apenas um caso de paralaxe cognitiva
» Nobel discorda da tese sobre mudanças climáticas
» Em busca de uma nova teoria que considere a complexidade biológica
» A complexidade tecnológica da celula da qual Darwin era totalmente ignorante
» E então a “tese” da gravidade criadora de Stephen Hawking era apenas um caso de paralaxe cognitiva
Permissões neste sub-fórum
Não podes responder a tópicos







 Seja fã Forumeiros
Seja fã Forumeiros
» Acordem adventistas...
» O que Vestir Para Ir à Igreja?
» Ir para o céu?
» Chat do Forum
» TV Novo Tempo...
» Lutas de MMA são usadas como estratégia por Igreja Evangélica para atrair mais fiéis
» Lew Wallace, autor do célebre livro «Ben-Hur», converteu-se quando o escrevia
» Ex-pastor evangélico é batizado no Pará
» Citações de Ellen White sobre a Vida em Outros Planetas Não Caídos em Pecado
» Viagem ao Sobrenatural - Roger Morneau
» As aparições de Jesus após sua morte não poderiam ter sido alucinações?