
 Início
InícioÚltimos assuntos
Tópicos mais visitados
Tópicos mais ativos
SEU IP


Michael Behe não foi refutado sobre o flagelo bacteriano
Página 1 de 1

29032012
Michael Behe não foi refutado sobre o flagelo bacteriano
“Minha queixa principal com os proponentes do Design Inteligente é que eles parecem nunca desistir. Quantas vezes alguém precisa lhes dizer que algo está errado antes que você admita? Quantas vezes o Design Inteligente precisa ser refutado na mídia com revisão por pares antes que você desista dela como uma causa perdida? A estória da complexidade irredutível do flagelo bacteriano está completa e totalmente morta. Está errada. Acostumem-se com isso.”
Recentemente, levantei a questão do flagelo bacteriano como sendo um exemplo documentado de complexidade irredutível numa sessão de perguntas e respostas num colóquio sobre a interseção da ciência e religião, e recebi respostas nesse mesmo sentido.
Mas essa afirmação é realmente verdadeira? Esse argumento tem sido refutado pelos críticos? Cerca de um ano atrás, li o livro Why Intelligent Design Fails - A Scientific Critique of the New Creationism [Por que o Design Inteligente falha - Uma crítica científica do neo-criacionismo] (editado por Matt Young e Taner Edis). O capítulo 5 desse livro foi contribuição de Ian Musgrave e tem como título “Evolution of the Bacterial Flagellum” [Evolução do flagelo bacteriano]. Direcionado como uma resposta a Michael Behe e William Dembski, Musgrave tenta, de uma vez por todas, demonstrar ser errada a noção de complexidade irredutível. Lendo esse capítulo, eu me lembro de ter ficado profundamente não impressionado. Na página 82 do livro, Musgrave nos oferece o seguinte argumento:
“Eis aqui um possível cenário [sic] para a evolução do flagelo bacteriano: primeiro surgiu um sistema de secreção, baseado ao redor do bastão SMC e do complexo de formação de poros, que foi o ancestral comum do sistema de secreção tipo III e do sistema flagelar. A associação de uma bomba de íon (que mais tarde se tornou a proteína do motor) a essa estrutura melhorou a secreção. Até hoje, as proteínas do motor, parte de uma família de proteínas que dirige a secreção, podem, livremente, desassociar-se e reassociar-se com a estrutura flagelar. O complexo do bastão e a formação de poros pode até ser rotacionado nesse estágio, como faz em alguns sistemas de deslizamento-mobilidade. O filamento protoflagelar surgiu em seguida como parte da estrutura de secreção de proteína (compare o Pseudomonas pilus, os apêndices filamentosos da Salmonella e as estruturas filamentosas da E. coli). A mobilidade em deslizar e contrair surgiu nesse estágio ou mais tarde, e depois foi refinada em mobilidade natatória. A regulação e a capacidade de manobrar podem ser adicionadas mais tarde, porque existem eubactérias modernas que não têm esses atributos, mas funcionam bem em seu ambiente (Shah e Sockett, 1995). Em cada estágio há um benefício para as mudanças na estrutura.”
Na verdade, Mark Pallen e Nick Matzke apresentaram argumento muito semelhante no artigo deles na Nature Reviews, em 2006 (artigo levantado por alguém no auditório durante o tour recente de Behe na Grã-Bretanha). Ken Miller também é reputado em fazer, rotineiramente, afirmações semelhantes concernentes à evolução do flagelo com o Sistema de Secreção Tipo III baseado largamente em considerações das homologias das sequências de proteínas.
Então, esses pontos tiveram sucesso em sepultar de uma vez por todas essa questão irritante do design inteligente? Bem, na verdade, não; eles não tiveram êxito. De fato, sugiro que os argumentos de todos esses cavalheiros há pouco mencionados trivializam fundamentalmente diversas questões importantes.
Primeiramente, trivializam a complexidade transparente e a sofisticação do sistema flagelar – tanto seu aparato de montagem quanto seu “motif” de design em seu nível mais alto de desenvolvimento – estado-da-arte. Na verdade, o processo de automontagem do flagelo bacteriano dentro da célula é tão sofisticado que tenho me esforçado há tempos para descrevê-lo de um modo acessível para leigos. Seus conceitos fundamentais são notoriamente difíceis de entender para aqueles que não estão acostumados a pensar sobre o sistema ou para aqueles que o estão encontrando pela primeira vez. Mas, ao mesmo tempo, a base mecanicista da montagem flagelar é tão elegantemente empolgante e mesmerizante que a transparente e esplêndida engenharia do motor flagelar – e, na verdade, a magnitude do desafio que ele traz para o darwinismo – não pode ser apreciada adequadamente sem um mínimo de conhecimento superficial de suas operações estruturais fundamentais. Vamos dar uma olhada.
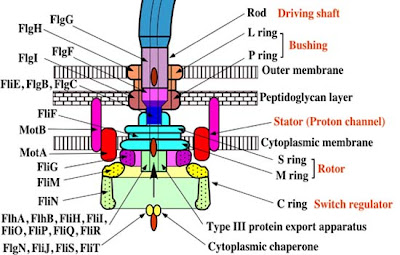
A síntese do flagelo bacteriano requer a expressão orquestrada de mais de 60 produtos de genes. Sua biossíntese dentro da célula é orquestrada por genes que são organizados em uma cascata bem ordenada na qual a expressão de um gene em um determinado nível exige a expressão anterior de outro gene em outro nível muito maior. O paradigma, ou modelo, de organismo para a montagem flagelar é a Salmonella, uma bactéria da família Enterobacteriaceae. Minha discussão, pois, pertence principalmente à Salmonella, a menos que seja indicada de outro modo.
O sistema flagelar na Salmonella tem três classes de promotores (os promotores são parecidos com um tipo de interruptor molecular que pode iniciar a expressão do gene quando reconhecido pelo RNA polimerase e uma proteína especializada associada chamada de “fator sigma”). Essas três classes de promotores são simplesmente chamadas de “Classe I”, “Classe II” e “Classe III”. Essa transcrição sequencial é acoplada ao processo de montagem flagelar. A Classe I contém apenas dois genes em um operon (chamado de FlhD e FlhC). A Classe II consiste de 35 genes ao longo de oito operons (inclusive genes envolvidos na montagem do corpo basal enganchado, e outros componentes do flagelo, bem como o aparato de exportação e dois genes reguladores chamados FliA e FlgM). Esses genes envolvidos na síntese do filamento são controlados pelos promotores da Classe III.

O promotor da Classe I dirige a expressão de um regulador mestre (particular às Enterobacteriaceae, da qual a Salmonella é membro) chamado FldH4C2 (não se preocupe se não se lembrar!). Esse regulador entérico mestre liga então os promotores da Classe II em associação com um fator sigma, σ70 (lembre de que eu disse que fator sigma é um tipo de proteína que capacita a união específica do RNA polimerase aos promotores de genes. Os promotores Classe II ficam então responsáveis pela expressão do gene das subunidades do corpo basal enganchado e seus reguladores, inclusive outro fator sigma chamado σ28 (que é codificado por um gene chamado FliA) e seus fatores antissigma, FlgM (os fatores antissigma, como seu nome sugere, se acoplam aos fatores sigma para inibir sua atividade transcricional). O fator sigma σ28 é exigido para ativar os promotores Classe III. Mas aqui nós, potencialmente, entramos num problema. Não faz, absolutamente, nenhum sentido começar a expressar os monômeros flagelinos antes de terminar a construção do corpo basal enganchado. Assim, a fim de inibir o σ28, o fator antissigma (FlgM) aludido acima inibe sua atividade e proíbe-o de interagir com o complexo holoenzimático da RNA polimerase. Quando a construção do corpo basal enganchado é completada, o fator antissigma FlgM é secretado através das estruturas flagelares que são produzidas pela expressão de genes de corpo basal enganchado Classe II. Os promotores da Classe III (que são responsáveis pela expressão dos monômeros flagelinos, do sistema de quimiotaxia e dos geradores de força motor) são finalmente ativados pelo σ28, e o flagelo pode ser completado.

Mas a coisa fica muito melhor. O sistema de exportação flagelar (isto é, o meio pelo qual o FlgM é removido da célula) tem dois estados substratos de especificidade: substratos tipo bastão/gancho e substratos tipo filamento. Durante o processo de montagem flagelar, esse interruptor de substrato-especificidade tem que se mover rapidamente daqueles estados anteriores para os últimos estados. As proteínas que formam parte do gancho e do bastão precisam ser exportadas antes que esses formem o filamento. Mas como surge esse interruptor no substrato-especificidade?
Uma proteína ligada à membrana chamada FlhB é a principal agente nesse processo. Há também uma proteína flagelar do tamanho do gancho que é responsável em se certificar de que o tamanho do gancho seja do tamanho correto (cerca de 55 nm) chamada FliK. Essa proteína também é responsável na ativação do interruptor de especificidade do substrato de exportação. Como se constata, sem a FliK, a capacidade de alternar e exportar filamento e o controle de tamanho do gancho são completamente perdidos. A FliK tem dois domínios principais, i.e. os domínios N-terminal e C-terminal. Durante a montagem do gancho, a FliKN funciona como um sensor molecular e transmissor de informação sobre o tamanho do gancho. Quando o gancho atinge o tamanho correto, a informação é transmitida à FliKC e à FliKCT, resultando numa mudança conformacional, que por sua vez resulta na ligação da FliKCT à FlhBC. Isso, por sua vez, resulta na mudança conformacional no FlhBC. E isso provoca a alternância do substrato de especificidade.
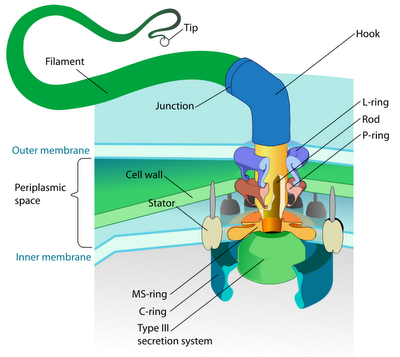
A montagem flagelar começa na membrana citoplásmica, avança pelo espaço periplásmico e, finalmente, se estende para fora da célula. Basicamente, o flagelo consiste de duas partes principais: o sistema de secreção e a estrutura axial. Os principais componentes da estrutura axial são a FlgG para o bastão, a FlgE para o gancho e a FliC para o filamento. Todas elas se reúnem com a assistência de uma proteína tampão (FlgJ, FlgD e FliD respectivamente). Dessas, somente a FliD permanece na ponta do filamento no produto final. Outros componentes da estrutura axial (chamados de FlgB, FlgC e FlgF) conectam o bastão ao complexo do anel MS. O gancho e o filamento são conectados pela FlgK e a FlgL.
Quando o anel C e o bastão C se acoplam ao anel M em sua superfície citoplásmica, o complexo do anel M – que é fundação estrutural do aparato – pode começar a secretar proteínas flagelares.
A estrutura do bastão é construída pela camada de peptidoglicano. Mas seu crescimento não é capaz de avançar além da barreira física apresentada pela membrana externa sem ajuda. Assim, o complexo do anel externo faz um buraco na membrana de modo que o gancho possa crescer por debaixo da base FlgD até que atinja o tamanho crítico de 55 nm. Então os substratos que estão sendo secretados podem alternar do modo bastão-gancho para o modo flagelino, o FlgD pode ser substituído por proteínas associadas ao gancho e o filamento continua a crescer. Sem a presença da proteína tampão FliD, esses monômeros flagelinos se perdem. Essa proteína tampão é essencial para que o processo ocorra.
Por que a evolução do flagelo do T3SS não funciona
Alguém pode ter pensado que a descrição dada acima seria mais do que suficiente para reduzir a nada os gestos de mão abanando das trivializações de Kenneth Miller et al. Mas fica cada vez pior para a estória darwinista. Por que a biossíntese do flagelo é tão precisamente regulada e orquestrada? Não somente as demandas de energia fazem do flagelo um sistema extremamente dispendioso para funcionar, mas a expressão inoportuna de proteínas flagelares pode induzir uma forte reação de imunização no sistema hospedeiro, algo que nenhuma bactéria quer sofrer.
Qual é o significado disso do ponto de vista da razão evolucionária? Bem, os monômeros flagelinos são algo tipo indutores de citoquinas. Se você fosse um organismo Yersinia, de posse de um Sistema de Secreção Tipo-III, a última coisa que você gostaria de fazer é apresentar esses peptídeos flagelinos aos macrofágos. Tal coisa, sem dúvida, seria prejudicial aos mecanismos anti-inflamatórios da Yersinia.
Conclusão
Minha descrição, dada acima, realmente somente tocou a superfície desse assunto espetacular de nanotecnologia (para mais detalhes, veja aqui). Por questão de brevidade, eu sequer discuti os processos impressionantes de quimiotaxia, dois componentes do circuito de transdução de sinal, o acoplamento rotacional, e a força motivo de próton pelo qual o flagelo é energizado (para detalhes disso, vide minha discussão aqui ou, para maiores detalhes, vide este artigo crítico). Mas a consideração mais importante é que a moderna teoria darwinista - como é classicamente entendida - não chegou nem perto de explicar a origem dessa máquina motor extraordinariamente complexa e sofisticada. Assim como as “explicações” darwinistas para o olho podem, a princípio, parecer convincentes para os não iniciados, grandemente não familiarizados com a absoluta maravilha de engenharia da bioquímica e da base molecular da visão, assim também as “explicações” evolucionárias do flagelo se tornaram rapidamente vazias de qualquer persuasão quando se consideram os detalhes moleculares do sistema. Quando alguém junta os detalhes acima com as demonstrações nítidas da impotência de o neodarwinismo produzir novas dobraduras de proteínas e novos sítios de ligação proteína-proteína, você pensa realmente que esse sistema pode ser ajuntado por virtude de leves, sucessivas modificações, um pequeno passo de cada vez? Considerando-se que o ponto importante de convencimento do neodarwinismo depende de sua suposta eficiência em invalidar a extraordinária aparência de design, não é óbvio que sua impotência demonstrável lança o postulado de design de volta à mesa como uma viável e respeitável proposição científica?
Douglas Axe, do Biologic Institute, demonstrou em artigo recente na revista Bio-complexity que o modelo de duplicação e recrutamento de gene somente funciona se algumas poucas mudanças forem necessárias para adquirir nova utilidade selecionável ou nova funcionalidade. Se um gene duplicado for neutro (em termos do seu custo para o organismo), então o número máximo de mutações que uma nova inovação numa população bacteriana pode exigir fica em torno de seis mutações. Se o gene duplicado tiver um custo adaptativo levemente negativo, o número máximo cai para dois ou menos (não incluindo a duplicação em si).
Parece que o flagelo bacteriano continua sendo - e talvez seja - um desafio muito maior ao darwinismo desde quando Behe escreveu o livro A Caixa Preta de Darwin, em 1996.
(Evolution News, via Desafiando a Nomenklatura Científica)
Michael Behe não foi refutado sobre o flagelo bacteriano

Eduardo- Mensagens : 5997
Idade : 54
Inscrição : 08/05/2010
Compartilhar este artigo em:
» Palestra de Michael Behe no Brasil sobre design inteligente
» O silêncio da Grande Mídia sobre Michael Behe no Brasil
» Flagelo bacteriano in situ: Darwin não explica a origem e nem a evolução disso!!!
» Entrevista com Michael Behe
» Michael Behe on Falsifying Intelligent Design (LEGENDADO).
» O silêncio da Grande Mídia sobre Michael Behe no Brasil
» Flagelo bacteriano in situ: Darwin não explica a origem e nem a evolução disso!!!
» Entrevista com Michael Behe
» Michael Behe on Falsifying Intelligent Design (LEGENDADO).
Permissões neste sub-fórum
Não podes responder a tópicos







 Seja fã Forumeiros
Seja fã Forumeiros
» Acordem adventistas...
» O que Vestir Para Ir à Igreja?
» Ir para o céu?
» Chat do Forum
» TV Novo Tempo...
» Lutas de MMA são usadas como estratégia por Igreja Evangélica para atrair mais fiéis
» Lew Wallace, autor do célebre livro «Ben-Hur», converteu-se quando o escrevia
» Ex-pastor evangélico é batizado no Pará
» Citações de Ellen White sobre a Vida em Outros Planetas Não Caídos em Pecado
» Viagem ao Sobrenatural - Roger Morneau
» As aparições de Jesus após sua morte não poderiam ter sido alucinações?